Abstract
Two new pleurothallid species of the genera Lepanthes and Pleurothallis, native to cloud forest remnants of the Eastern Andes of Colombia, are described and illustrated. Lepanthes pseudoabitaguae is compared with Lepanthes abitaguae, from which it differs by the color of the flowers and by the oblong and biglandular appendix. Pleurothallis falcata is similar to Pleurothallis suspensa, but can be recognized by the falcate to lanceolate leaves, denser inflorescences, longer sepals, and the lip completely recurved with erose apical margins. We also provide comparisons with similar species and offer comments on their ecology. Furthermore, we discuss the significance of this highly anthropized region and its richness in orchid species.
Keywords: endemism; flora of Colombia; Lepanthes; Pleurothallis; taxonomy
Resumen
Se describen e ilustran dos nuevas especies de Pleurothallidinae de los géneros Lepanthes y Pleurothallis, nativas de remanentes de bosque nublado en los Andes Orientales de Colombia. Lepanthes pseudoabitaguae se compara con Lepanthes abitaguae, de la cual difiere por el color de las flores y por el apéndice oblongo y biglandular. Pleurothallis falcata es similar a Pleurothallis suspensa, pero puede ser reconocida por las hojas falcadas a lanceoladas, inflorescencias más densas, sépalos más largos y el labelo completamente recurvo con márgenes apicales erosas. También proporcionamos comparaciones con especies similares y ofrecemos comentarios sobre su ecología. Además, discutimos la importancia de esta región altamente antropizada y su riqueza en especies de orquídeas.
Palabras clave: endemismo; flora de Colombia; Lepanthes; Pleurothallis; taxonomía
Introduction
The tropical montane cloud forests constitute nearly 2.5% of the surface of tropical forests. Despite their relatively small extent, they are of utmost priority for biodiversity conservation, considering the outstanding values of endemic and threatened species they harbor (Armenteras et al. 2007, Bubb et al. 2004). Moreover, these forests play a crucial role in providing fresh water and storing carbon, among other benefits for human well-being (Aparecido et al. 2018).
Neotropical cloud forests exhibit exceptional diversity, particularly in certain groups of birds, amphibians, and vascular epiphytes, notably orchids (Carmona-Higuita et al. 2023, Gentry & Dodson 1987, Pérez-Escobar et al. 2017). Among orchids, the Pleurothallidinae subtribe show one of the highest evolutionary diversification rates, representing a significant portion of the orchid flora found in montane forests (Pérez-Escobar et al. 2017). A recently published catalogue of the Pleurothallidinae subtribe for Colombia listed 1854 species, with almost 60% of them being endemic to the country. The most species richn genera are Stelis Sw. (521 species), Lepanthes Sw. (377 species) and Pleurothallis R.Br. (248 species) (Karrremans et al. 2023).
In the Neotropics, the major threat to this unique ecosystem is the clearance for cattle ranching. In the case of Colombia, despite trends of rural depopulation, the expansion of grasslands has continued (Bubb et al. 2004), resulting in cloud forests becoming isolated islands on mountain tops or along steep hills, remnants of the once extensive areas of Andean forests. The eastern range of the Colombian Andes, known as “Cordillera Oriental,” has undergone extensive transformation, particularly towards the Magdalena Valley. Approximately 51% of its original natural ecosystems have been lost, with only 45% of the original cover of Andean forests remaining, and less than 5% designated as protected areas, as estimated two decades ago (Armenteras et al. 2003).
However, small remnants of well-preserved cloud forests within transformed landscapes play a crucial role as a refuge for epiphyte diversity (Koster et al. 2009). This significance increases when considering narrow endemic species such as orchids of the Pleurothallidinae subtribe.
Previously unexplored areas of the Neotropics may host several undescribed plant species (Gentry 1982). Recent studies of the orchid flora in remnants of cloud forests in a floristically rich area of the eastern Andes in Santander, Colombia, have led to the discovery of new orchid species (Gutiérrez Morales et al. 2018, 2021, 2023, Hágsater et al. 2018, Moreno et al. 2018, Vieira-Uribe & Gutiérrez 2020). As part of this ongoing research, we present two new orchid species from the same locality, specifically of the genera Lepanthes and Pleurothallis within the Pleurothallidinae subtribe.
Taxonomic treatment
Lepanthes pseudoabitaguae N.Gut., K.Gil-Amaya & J.S.Moreno sp. nov. (Fig. 1, 2, 3A)


TYPE: COLOMBIA. Santander: La Belleza, Vereda El Chircal-Ojo de Agua, 2530 m. 18 November 2016, N. Gutiérrez 17 (holotype, JBB 17269).
Diagnosis: Lepanthes pseudoabitaguae is most similar to Lepanthes abitaguae Luer & L.Jost, as its name suggests. The primary distinguishing feature lies in their appendices which in L. abitaguae is triangular, positioned below the stigma and acutely reflexed (vs. oblong and biglandular in L. pseudoabitaguae).
Plant epiphytic, caespitose, up to 22 cm tall. Roots slender, flexuous, filiform, ca. 1.0-1.2 mm in diameter. Ramicauls slender, suberect, 3.5-14 cm long, enclosed by 7-13 ribbed, minutely ciliate, acuminate, lepanthiform sheaths. Leaves suberect, papyraceous, purple on the abaxial surface, lustrous, likely iridescent, ovate to elliptical, the margins recurved, reticulate-veined, obtuse, the apex emarginate with an abaxial central apiculum, 4.5-7.2 cm × 1.7-2.3 cm wide, the base cuneate, contracted into a petiole 1.0-1.2 mm long. Inflorescence (1-2) per ramicaul, a congested, distichous raceme successively many-flowered, up to 4.5 cm long, borne on the adaxial side of the leaf by a slender and filiform peduncle, 2.0-3.7 cm long; floral bracts conical, acuminate, 1.4-1.9 mm long. Ovary costate, 3.3-3.9 mm long. Flowers with sepals magenta to pale red, with pale yellow at the margins, petals and lip cinnabar, the column vermilion with the apex claret, the anther cap purple. Dorsal sepal glabrous, ovate, slightly concave in the middle, acute-attenuate, 3-veined, 5.0-6.7 × 3.8-5.5 mm, connate to the lateral sepals for up to 2.65 mm. Lateral sepals shortly papillate along the margins, ovate, oblique, each-2-veined, 4.4-6.1 × 2.4-3.3 mm, connate up to 3.4 mm into an ovate, bifid synsepal with attenuate, divergent apices. Petals transversally trilobed, 3-veined, minutely pubescent, the upper lobe quadrate, oblique, 4 × 2 mm, 2 mm long, the flagelliform midlobe uncinate from the outer margin between the lobes, the lower lobe narrowly triangular, acute, 3.7 × 1.2 mm. Lip bilaminate, the blades suboblong, 1.7-2.6 × 0.6-0.9 mm, the base obtuse, the apex truncate, minutely pubescent, sharply folded over the thin and broadly cuneate connectives, adnate to the base of the column, with a pubescent, oblong, sinuose, biglandular appendix. Column terete, stigma ventral, bilobed, anther dorsal, elongated beyond the stigma, 2.9 mm long. Anther cap cucullate, elongate, 1.7 × 0.5 mm. Pollinia 2, yellow, narrowly oblanceolate, attached to a viscidium, 1.4 mm long.
Additional material examined: COLOMBIA. Santander: La Belleza, Vereda El Chircal-Ojo de Agua, 2583 m. 11 May 2022, N. Gutiérrez 232, (paratype, JBB). Bolivar, Hoya de Panamá, 2790 m. February 2016 (photo voucher: David Haelterman).
Eponymy: Named due to its remarkable similarity with Lepanthes abitaguae.
Distribution: Lepanthes pseudoabitaguae is apparently restricted to montane humid forests on the western slope of the Eastern Andes of Colombia, in La Belleza Municipality, Santander (Fig. 4). Despite the high degree of fragmentation of these cloud forests, this species has been observed several times in the type locality and once in a second area located near the top of the “Hoya de Panamá,” in the Municipality of Bolivar, Santander.
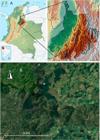
A. Santander department in the Eastern Andes of Colombia, the specific region is marked to the south.
Habitat and ecology: The type specimen was found growing near the ground on branches densely covered by bryophytes, with limited exposure to light. Other Lepanthes species registered in these forest remnants include Lepanthes agglutinata Luer, Lepanthes saccata Luer & R.Escobar, Lepanthes palaga Luer & R.Escobar and Lepanthes dryades Luer & R.Escobar.
Discussion
The most similar species to Lepanthes pseudoabitaguae is undoubtedly Lepanthes abitaguae (Fig. 3B), endemic to the eastern slope of the Andes of Ecuador. The plants are similar in overall size and in the proportions between ramicauls and leaves, the leaves are iridescent, suffused with purple beneath. The flowers are apparently identical, just differing in the color of petals and sepals (yellow-orange in L. abitaguae, magenta to pale red in L. pseudoabitaguae), in the degree of connation of the dorsal sepal to the lateral sepals (1 mm in L. abitaguae, 3.4 mm in L. pseudoabitaguae), and in the appendix morphology.
When we first collected and photographed the new species, we erroneously identified it as Lepanthes abitaguae due to their remarkable similarity in plant and flower morphology. However, considering the distribution range of L. abitaguae, and some minor discrepances in flower morphology that we mentioned earlier, we decided to dissect the specimens under a stereo microscope to confirm its identity. Focusing on the defining feature of many Lepanthes species, which is the appendix, we found a significant difference in this structure, which is oblong and biglandular in Lepanthes pseudoabitaguae, but reflexed and triangular in L. abitaguae. This is the most distinct difference between the two species.
On the other hand, it is worth noting that when describing L. abitaguae, Luer did not provide detailed descriptions or illustrations of the column, stigma, or anther, which may also vary between both species. Instead, he placed emphasis on the lip blades and the appendix.
Pleurothallis falcata N.Gut. & K.Gil-Amaya, sp. nov. (Fig. 5, 6, 7A-B)



TYPE: COLOMBIA Santander: La Belleza, Vereda El Chircal-Ojo de Agua, 2457 m. 14 January 2023, N. Gutiérrez 270 (holotype, JBB 38541; isotype, N. Gutiérrez 271, JBB 38542).
Diagnosis: Similar to Pleurothallis suspensa Luer, distinguished by the falcate to lanceolate leaves (vs. elliptical-ovate), the denser inflorescences with pinkish and pale yellow flowers (vs. inflorescences with distant, purple flowers), the larger sepals, (sepals >10.5 mm vs.<8.0 mm long), and the lip with the apex completely recurved, with the apiculum recurved and the apical margins erose (vs. lip partially recurved, the apiculum straight, the apical margins entire).
Plant epiphytic, caespitose, up to 27 cm tall. Roots slender, flexuous, ca. 1 mm in diameter. Ramicauls erect to suberect, slender, 8.5-16.5 cm long, partially enclosed by a pair of close, tubular, ribbed, truncate sheaths. Leaf suberect, thinly coriaceous, falcate to lanceolate, the apex attenuate, acuminate, 6.3-11 × 1.0-1.3 cm, the base cuneate. Inflorescence 1-10 per ramicaul, loose, pendant, fragile, subcongested, 5-10 flowered racemes of simultaneous, pinkish to light yellow, non-resupinate flowers, 6.0-9.5 cm long including the peduncle ca. 5 cm long, emerging from a spathe ca. 1-2 cm long at the base of the leaf; floral bracts tubular, slightly dilated, truncate, 4 mm long, pedicels 3 mm long. Ovary green to vinaceous, sulcate, 2 mm long. Flowers not resupinate, pinkish to pale yellow. Sepals membranous, glabrous, the dorsal sepal narrowly lanceolate, acute, 9.7 × 1.9 mm, 3-veined, the lateral sepals completely connate into a lanceolate, deeply concave, acute synsepal, 10.5 × 2.8 mm, each 2-veined. Petals membranous, narrowly linear, acute, 8.6 × 0.4 mm, 3-veined. Lip vinaceous, darker centrally, broadly ovate, 3-lobed, arcuate, 1.6 × 1.7 mm unexpanded, the lateral lobes erect, broadly rounded, slightly oblique, the midlobe with the apex apiculate, erose, vinaceous, the disc convex, slightly sulcate, without callus, the base truncate, hinged to the column-foot. Column stout, semiterete, 1.2 mm long. The anther and stigma apical, the rostellum elongate, triangular, tinged with rose. Pollinia two, yellow, ovoid, ca. 0.4 mm long, attached to a rounded viscidium. Fruit 0.8 cm long, 0.6 cm wide.
Additional material examined: COLOMBIA. Santander: La Belleza, Vereda El Chircal-Ojo de Agua, 2530 m. 31 May 2017, N. Gutiérrez 63 & S. Moreno (paratype, JBB 17607).
Etymology: From the Latin falcatus, “hooked, curved like a scythe or sickle,” in allusion to some of the leaves of this species, such character is more evident in the photograph of the paratype specimen (Fig. 7A).
Distribution: Only known from the western slope of the Eastern Andes, in La Belleza, Santander, Colombia, where it inhabits cloud forests at elevations of 2400 to 2600 m, in the same forest remnants that host populations of Lepanthes pseudoabitaguae.
Habitat and ecology: Epiphytic in partial shade in mature vegetation, on trunks of several Melastomataceae trees and tree ferns of the genus Cyathea Sm. (Cyatheaceae). The type specimen was found alongside other orchid species such as Lepanthes palaga, L. saccata, and Oliveriana ortizii Á.Fernández. Flowering occurs from January to May. Individuals from different populations exhibit variations in overall flower size and color, ranging from entirely white, cream to pink, while the leaves of a single specimen may also display varying degrees of curvature.
Discussion. Pleurothallis falcata can be distinguished by the elongated ramicauls with falcate to lanceolate leaves, pendant, loose racemes of pinkish or yellow, non-resupinate flowers, the synsepal deeply concave, the narrow-linear petals and the recurved lip with prominent lateral lobes, with the apex apiculate and minutely erose. Pleurothallis falcata is similar to some species included in Luer’s (1999) revision of Pleurothallis, subgen. Pleurothallis, subsection Antennifera Luer, such as Pleurothallis suspensa, which occurs in lowlands of Suriname, Guyana, and Venezuela; and Pleurothallis antennifera Lindl., widely distributed in the mid to high altitudes in the Andes of Venezuela to Bolivia.
Pleurothallis suspensa is without a doubt the most similar species, because of its overall size of the plant, with slender, elongated ramicauls, loose, pendant racemes with no resupinate, simultaneous flowers, the petals narrowly linear, and the recurved lip with the sides broadly rounded, the disc slightly sulcate and the apex minutely apiculate. Pleurothallis falcata differs by the longer, lanceolate to falcate leaves (vs. elliptical-ovate leaves), the congested racemes (vs. racemes with flowers distant from each other), the flowers pinkish or pale yellow (vs. flowers purple), the sepals larger, the dorsal sepal narrowly lanceolate (vs. narrowly ovate), the deeply concave synsepal (vs. concave), and the lip with the apex completely recurved, with the apiculum recurved and the apical margins erose (vs. lip partially recurved, the apiculum straight, the apical margins entire).
Pleurothallis antennifera is also similar to the new species, albeit to a lesser extent. It shares the elongated ramicauls, lanceolate leaves, subcongested inflorescences with simultaneous flowers, linear petals, concave synsepal, lip concave with pronounced lateral lobes, with the apical margins erose and apiculate at the apex. It can be easily distinguished from P. falcata by the larger size, reptant habit (vs. caespitose habit), the absence of falcate leaves, the erect inflorescences (vs. pendant), the shape of the sepals, and the lip with the lateral lobes oblong, incurved, and the apex incurved (vs. lobes rounded, straight, apex recurved).
The new species can be treated within Pleurothallis subsection Antennifera considering the elongate leaves, narrowly linear petals, recurved lip with prominent and broad lateral lobes, and the undeveloped glenion. However, contrary to Luer´s (1999) statement that erect inflorescences with resupinate flowers are typically frequent in subsection Antennifera, the pendent raceme of Pleurothallis falcata carries no resupinate flowers. Such a combination of characters seems uncommon within the subsection Antennifera but is commonly found in subsect Macrophyllae-Racemosae Luer.
It’s important to emphasize that the subsection Antennifera, as noted by Wilson et al. (2017), is not a natural group, instead, it is embedded within a larger group that also includes the species of Pleurothallis subsect. Macrophyllae-Racemosae.
Final considerations
With the discovery of these two new taxa, we have identified seven new species from five orchid genera in this area of fragmented cloud forest. Interestingly, all the species are within 5 km from each other, within an elevational range of 2350 to 2570 m (Fig. 4B, 7C-D). Our comparisons of the orchid community between fragments indicates a high species turnover (Gutiérrez Morales unpubl.). For example, there is no more than 2.6 km between the three Lepanthes species described, Lepanthes agatarum J.S.Moreno, N.Gut. & S.Vieira-Uribe, Lepanthes marielana N.Gut., J.S.Moreno & S.Vieira-Uribe, and Lepanthes pseudoabitaguae. To date, they have not been seen growing close or together.
Furthermore, the inventory of the orchid species in this specific area has also contributed several new records to the Colombian flora: Andinia ciliaris (Luer & Hirtz) Karremans & S.Vieira-Uribe, Lankesteriana caudatipetala (C.Schweinf.) Karremans, Platystele jesupiorum Luer, and Ophidion compactum (Vierling) Karremans & J.S.Moreno, reported in Karremans et al. (2023). Additionally, Myoxanthus parvilabius (C.Schweinf.) Luer, as reported in Rojas Alvarado et al. (2021).
Unfortunately, the specific region of the Eastern Andes in Colombia lacks adequate protected areas. Consequently, the outstanding vegetal diversity in this area faces a high level of threat and can gradually disappear due to the ongoing expansion of grasslands and selective logging (Fig. 7C-D). The absence of protective measures puts these unique species at risk of gradual disappearance. Therefore, the establishment of natural reserves, even if they are small in size, is imperative for the long-term conservation of these species and the preservation of the still undiscovered biodiversity within these forests
Acknowledgements
We would like to thank the José Celestino Mutis Botanical Garden in Bogotá for the collection permits, with special thanks to Boris Villanueva, and the entire staff of the herbarium who assisted with the preparation of voucher specimens. Our sincere appreciation also goes to Fernando Morales, uncle of the first author, for bringing us to the remnant cloud forests where the two new species were discovered. We also thank the anonymous reviewers for their valuable comments on the manuscript.
Literature cited
- Aparecido, L. M. T., Teodoro, G. S., Mosquera, G., Brum, M., Barros, F., Pompeu, P. V., Rodas, M., Lazo, P., Muller, C. S., Mulligan, M., Asbjornsen, H, Georgianne W. Moore, G.W. & Oliveira, R. S. (2018). Ecohydrological drivers of Neotropical vegetation in montane ecosystems. Ecohydrology, 11(3), e1932.
- Armenteras, D., Gast, F. & Villareal, H. (2003). Andean forest fragmentation and the representativeness of protected natural areas in the eastern Andes, Colombia. Biological conservation 113, 245-256.
- Armenteras, D., Cadena-V, C., Moreno, R. P. (2007). Evaluación del estado de los bosques de niebla y la meta 2010 en Colombia Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá. D.C, Colombia, 72p.
- Bubb, P., May, I. A., Miles, L. & Sayer, J. (2004). Cloud forest agenda Cambridge, UK: UNEP World Conservation Monitoring Centre.
-
Carmona-Higuita, M. J., Mendieta-Leiva, G., Gómez-Díaz, J. A., Villalobos, F., Ramos, F. N., Elias, J. P. C., Jiménez-López, D. A., Zuluaga, A., Holst, B., Kressler, M., Mathieu, G., Zizka, A., Zotz, G. & Krömer, T. (2023). Conservation status of vascular epiphytes in the Neotropics. Research Square, https://doi.org/10.21203/rs.3.rs-2773328/v1.
» https://doi.org/https://doi.org/10.21203/rs.3.rs-2773328/v1 - Gentry, A. H. (1982). Neotropical floristic diversity: phytogeographical connections between Central and South America, Pleistocene climatic fluctuations, or an accident of the Andean orogeny? Annals of the Missouri Botanical Garden 69, 557-593.
- Gentry, A. H., & Dodson, C. H. (1987). Diversity and biogeography of neotropical vascular epiphytes. Annals of the Missouri Botanical Garden, 74(2), 205-233.
- Gutiérrez Morales, N., Harding, P., Dalström, S. & Moreno, J. S. (2018). A new species of Cyrtochilum (Orchidaceae: Oncidiinae) from the Eastern Cordillera of Colombia. Lankesteriana 18(1), 63-68.
- Gutiérrez Morales, N., Moreno, J. S., Karremans, A. P. & Gil-Amaya, K. (2023). Restrepia santanderensis (Orchidaceae: Pleurothallidinae), a new species from the western slope of the eastern Andes in Colombia. Phytotaxa, 598(4), 293-300.
- Gutiérrez Morales, N., Vieira-Uribe, S. & Moreno, J. S. (2021). Lepanthes marielana (Orchidaceae, Pleurothallidinae), a new species from the eastern Andes in Colombia. Phytotaxa, 484(1), 113-120.
- Hágsater, E., Gutiérrez Morales, N. & Santiago, E. (2018). Epidendrum moralesii In Hágsater, E. & Santiago, E. (Eds.) The Genus Epidendrum, Part 12, Icones Orchidacearum 16 (1). Instituto Chinoin, A. C., México, t. 1646.
- Karremans, A. P., Moreno, J. S., Gil-Amaya, K., Gutiérrez Morales, N., Espinosa, F., Mesa, S., Restrepo, E., Rincón-González, M., Serna, A., Sierra-Ariza, M & Vieira-Uribe, S. (2023). Colombian Orchidaceae: A catalogue of the Pleurothallidinae. Lankesteriana, 23(2), 181-400.
-
Köster, N., Friedrich, K., Nieder, J., Barthlott, W. (2009). Conservation of epiphyte diversity in an Andean landscape transformed by human land use. Conservation Biology, 23(4), 911-919. https://doi.org/10.1111/j.1523-1739.2008.01164.x
» https://doi.org/https://doi.org/10.1111/j.1523-1739.2008.01164.x - Luer, C. A. (1999). Icones Pleurothallidinarum XVIII. Systematics of Pleurothallis, subgen. Pleurothallis, Sect. Pleurothallis, Subsect. Antenniferae, Subsect. Longiracemosae, Subsect. Macrophyllae-Racemosae, Subsect. Perplexae, Subgen. Pseudostelis, Subgen. Acuminatia Addenda to Dracula, Lepanthes, Masdevallia and Pleurothallis Miscellaneous new species of Dryadella, Lepanthes and Pleurothallis Monographs in Systematic Botany from the Missouri Botanical Garden 76: 1-182.
- Moreno, J. S., Gutiérrez Morales, N. & Vieira-Uribe, S. (2018). Lepanthes agatarum (Orchidaceae: Pleurothallidinae), a new species honoring an ancient tribe from the eastern Andes in Colombia. Phytotaxa, 376(2), 89-96.
-
Pérez-Escobar, O. A., Chomicki, G., Condamine, F. L., Karremans, A. P., Bogarín, D., Matzke, N. J., Silvestro, D., Antonelli, A. (2017). Recent origin and rapid speciation of Neotropical orchids in the world’s richest plant biodiversity hotspot. New Phytologist 215(2), 891-905. https://doi.org/10.1111/nph.14629
» https://doi.org/https://doi.org/10.1111/nph.14629 - Rojas-Alvarado, G., Blanco, M. A. & Karremans, A. P. (2021). A taxonomic synopsis and morphological characterization of Myoxanthus (Orchidaceae: Pleurothallidinae). Phytotaxa, 507(3), 211-258.
- Vieira-Uribe, S. & Gutiérrez Morales, N. (2020). Andinia auriculipetala, sp. nov. Species Orchidacearum 4(1), LCDP 37.
- Wilson, M., Dupree, K., Garcia Lopera, D., Haelterman, D., Kay, A., Mesa Londoño, C., Niessen, A., Pinnix, W., Portilla, J. & Werner, J. D. (2017). A new species of Pleurothallis (Orchidaceae: Pleurothallidinae) from Valle del Cauca, Colombia and a note on the relationship between subsections Macrophyllae-Racemosae and Antenniferae Lankesteriana, 17(2), 119-131.
Publication Dates
-
Date of issue
Jan-Apr 2024
History
-
Received
11 Oct 2023 -
Accepted
23 Dec 2023

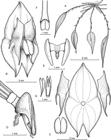
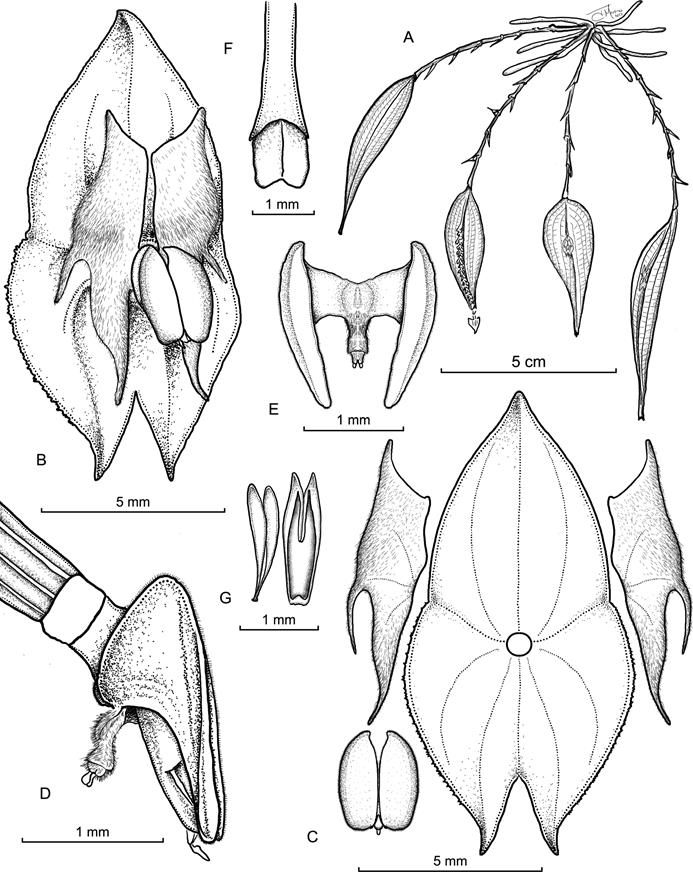 Illustration by Juan Sebastián Moreno based on the paratype.A. Habit. B. Flower. C. Perianth, with lateral view of the synsepal. D. Ovary, column and lip, lateral view. E. Lip, ventral view. F. Column, dorsal view. G. Anther cap and pollinia.
Illustration by Juan Sebastián Moreno based on the paratype.A. Habit. B. Flower. C. Perianth, with lateral view of the synsepal. D. Ovary, column and lip, lateral view. E. Lip, ventral view. F. Column, dorsal view. G. Anther cap and pollinia.
 Photos by Juan Sebastián Moreno.A. Flower, frontal view. B. Flower, lateral view. C. Habit.
Photos by Juan Sebastián Moreno.A. Flower, frontal view. B. Flower, lateral view. C. Habit.
 Photos by Juan Sebastián Moreno (A) and Andreas Kay (B).A. Lepanthes pseudoabitaguae.B. Lepanthes abitaguae.
Photos by Juan Sebastián Moreno (A) and Andreas Kay (B).A. Lepanthes pseudoabitaguae.B. Lepanthes abitaguae.
 Photos by Wikipedia (A) and Google Earth (B).B. Satellite images of the forest fragments that harbors the two new species, along with several other endemic orchid species, within a highly transformed landscape.
Photos by Wikipedia (A) and Google Earth (B).B. Satellite images of the forest fragments that harbors the two new species, along with several other endemic orchid species, within a highly transformed landscape.
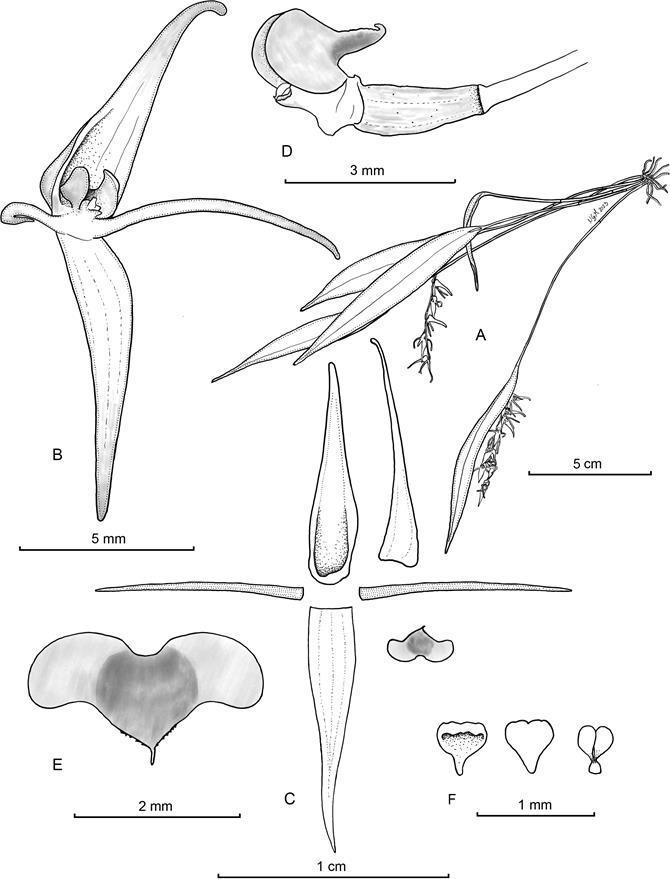 Illustration by Nicolás Gutiérrez Morales based on the paratype.A. Habit. B. Flower. C. Perianth, with lateral view of the synsepal. D. Ovary, column and lip, lateral view. E. Inverted, expanded lip. F. Anther cap and pollinarium.
Illustration by Nicolás Gutiérrez Morales based on the paratype.A. Habit. B. Flower. C. Perianth, with lateral view of the synsepal. D. Ovary, column and lip, lateral view. E. Inverted, expanded lip. F. Anther cap and pollinarium.
 Photos by Nicolás Gutiérrez Morales.A. Habit. B. Flower. C. Periant. D. Ovary, column and lip, lateral view. E. Lip, dorsal and lateral views. F. Column, front view. G. Column, ventral view. H. Anther and pollinarium.
Photos by Nicolás Gutiérrez Morales.A. Habit. B. Flower. C. Periant. D. Ovary, column and lip, lateral view. E. Lip, dorsal and lateral views. F. Column, front view. G. Column, ventral view. H. Anther and pollinarium.
 Photos by Nicolás Gutiérrez Morales (B-D), and courtesy of the JBB (A).B. Pleurothallis falcata in situ. C. Clearance of cloud forest to grasslands. D. Forest remnants in steep hills.
Photos by Nicolás Gutiérrez Morales (B-D), and courtesy of the JBB (A).B. Pleurothallis falcata in situ. C. Clearance of cloud forest to grasslands. D. Forest remnants in steep hills.