Abstract
The fourth release of the series “New Species and Records of Orchidaceae from Costa Rica” documents and illustrates 17 taxa of the Costa Rican orchid flora. The new species records belong to the subtribes Pleurothallidinae (9 spp.), Maxillariinae (3 spp.), and Oncidiinae (2 spp.), including the description of two new species in Pleurothallis and Telipogon. Two new forms are described for Isochilus latibracteatus (Ponerinae) and Masdevallia striatella (Pleurothallidinae), and their differences from the typical forms are discussed. Additionally, the first record of a naturalized population of Phalaenopsis stuartiana (Aeridinae) in Costa Rica is discussed. Detailed descriptions, based on selected Costa Rican material, are included for all taxa, along with illustrations or Lankester Composite Digital Plates (LCDP). Information on their etymology, distribution, habitat, phenology, and distinguishing features compared to morphologically similar species is also provided. Finally, following the publication of the most recent Costa Rican orchid catalogue, we have identified some omissions and other new species described, which are discussed here. The orchid flora of Costa Rica now includes 1695 species and nine forms.
Keywords: Aeridinae; Costa Rican orchids; Flora Costaricensis; Maxillariinae; Oncidiinae; Pleurothallidinae
Resumen
La cuarta entrega de la serie “Nuevas especies y registros de Orchidaceae de Costa Rica” documenta e ilustra 17 taxones para la flora de orquídeas costarricenses. Los nuevos registros de especies pertenecen a las subtribus Pleurothallidinae (9 spp.), Maxillariinae (3 spp.) y Oncidiinae (2 spp.), incluyendo la descripción de dos especies nuevas en Pleurothallis y Telipogon. Se describen dos nuevas formas para Isochilus latibracteatus (Ponerinae) y Masdevallia striatella (Pleurothallidinae), y se discuten sus diferencias con las formas típicas. Además, se discute el primer registro de una población naturalizada de Phalaenopsis stuartiana (Aeridinae) en Costa Rica. Se incluyen descripciones detalladas, basadas en material costarricense seleccionado, para todas las especies, junto con ilustraciones o láminas digitales compuestas Lankester (LCDP). También se proporciona información sobre su etimología, distribución, hábitat, fenología y características distintivas comparadas con especies morfológicamente similares. Finalmente, tras la publicación del más reciente catálogo de orquídeas de Costa Rica, hemos identificado algunas omisiones y otras nuevas especies descritas que se comentan aquí. La flora de orquídeas de Costa Rica incluye ahora 1695 especies y nueve formas.
Palabras clave: Aeridinae; Flora Costaricensis; Maxillariinae; Oncidiinae; orquídeas de Costa Rica; Pleurothallidinae
Introduction
In 2003, Robert L. Dressler presented the most comprehensive treatment of Orchidaceae available to date for Costa Rica (Dressler 2003). Subsequently, the series “New Species and Records of Orchidaceae from Costa Rica” was initiated to formally document newly discovered species and report those previously unknown in the country. Three contributions to this series were published between 2008 and 2014 (Bogarín et al. 2008, Fernández et al. 2014, Karremans et al. 2012). Since the latest publication, 114 orchid species from Costa Rica have been described as new to science, averaging approximately 11 new species per year. These belong to multiple diverse genera, such as Anathallis Barb.Rodr. (Karremans & Vieira-Uribe 2020), Andreettaea Luer (= Muscarella Luer) (Fernández et al. 2021), Brachionidium Lindl. (Bogarín et al. 2015, Bogarín & Karremans 2016), Campylocentrum Benth. (Bogarín 2015), Cischweinfia Dressler & N.H.Williams (Pupulin et al. 2020a), Crossoglossa Dressler & Dodson (Ormerod 2014), Daiotyla Dressler (Pupulin 2019), Dichaea Lindl. (Pupulin 2019, Pupulin & Karremans 2020), Echinosepala Pridgeon & M.W.Chase (Pupulin et al. 2017c, 2020b, 2022), Epidendrum L. (Díaz-Morales & Karremans 2016, Karremans 2021), Eurystyles Wawra (Bogarín 2020), Lepanthes Sw. (Bogarín et al. 2018a, 2019, 2020, Bogarín & Kisel 2014, Chinchilla et al. 2020, Larsen 2014, Pupulin 2020, 2021, Pupulin & Bogarín 2014a,b), Lockhartia Hook. (Blanco 2014), Malaxis Sol. ex Sw. (Chinchilla et al. 2022, 2023), Masdevallia Ruiz & Pav. (= Reichantha Luer) (Bogarín et al. 2017, Smith et al. 2015, Pupulin 2020), Maxillariella M.A.Blanco & Carnevali (Bogarín et al. 2018b), Mormodes Lindl. (Blanco et al. 2016), Myoxanthus Poepp. & Endl. (Rojas-Alvarado & Karremans 2017), Oncidium Sw. (Pupulin 2020), Pelexia Poit. ex Lindl. (Bogarín & Pupulin 2021), Platystele Schltr. (Karremans & Bogarín 2017), Pleurothallis R.Br. (Karremans & Jiménez 2018, 2023, Pupulin 2020, Pupulin et al. 2017a,b, 2021), Polycycnis Rchb.f. (Gerlach & Pupulin 2020), Prescottia Lindl. (Pupulin 2020), Prosthechea Knowles & Westc. (Bogarín & Karermans 2015), Pterichis Lindl. (Kolanowska 2014), Sarcoglottis C.Presl (Bogarín & Pupulin 2021), Scaphosepalum Pfitzer (Karremans & Pupulin 2022), Sobralia Ruiz & Pav. (Dressler & Pupulin 2014, 2015, Dressler et al. 2014, 2016, Pupulin & Díaz-Morales 2022), Specklinia Lindl. (Bogarín et al. 2014, Karremans & Vieira-Uribe 2020, Karremans et al. 2015a,b, 2020), Stelis Sw. (Bogarín & Pupulin 2019, Karremans & Díaz-Morales 2017, Karremans & Rojas-Alvarado 2022, Rojas-Alvarado 2023), Telipogon Kunth (Bogarín et al. 2024, Pupulin & Bogarín 2022), Vanilla Mill. (Karremans & Lehmann 2018), and Zootrophion Luer (Pupulin & Rojas-Alvarado 2022). Also, new nothospecies have been proposed in Cochlezella J.M.H.Shaw, Pleurothallis, and Phragmipedium Rolfe (Pupulin 2015, Pupulin & Díaz-Morales 2018, Pupulin et al. 2021).
The most updated comprehensive catalogue of the Costa Rican orchid flora (Pupulin et al. 2023) accounts for 1684 species, four subspecies, 14 natural hybrids, and seven forms across 201 genera and two nothogenera. Here, we present detailed descriptions, illustrations, or Lankester Composite Digital Plates (LCDP) for 12 new records previously listed in Pupulin et al. (2023) and two new species to science in the Oncidiinae and Pleurothallidinae. Additionally, we formally describe and illustrate two new forms for Isochilus latibracteatus A.Rich. & Galeotti (Ponerinae) and Masdevallia striatella Rchb.f. (Pleurothallidinae). Furthermore, we discuss the first record of a population of Phalaenopsis stuartiana Rchb.f. (subtribe Aeridinae) naturalized in Costa Rica. Along with the generic monographs and taxonomic treatments for groups of closely related orchids in larger genera, this series represents our ongoing effort to contribute to Flora Costaricensis, with the aim of providing a comprehensive treatment of Costa Rican orchids (Atwood & Mora de Retana 1999, Bogarín et al. 2014, Pupulin 2010, Pupulin & Bogarín 2010, 2014a,b, Pupulin et al. 2020b, 2021, Rojas-Alvarado & Karremans 2020).
Materials and methods
Plants collected in Costa Rica between March 2002 and October 2023, were labeled and cultivated at the Lankester Botanical Garden (JBL), where their study and documentation were carried out. Field notes for each collection included geographical, phenological, and taxonomic data. At blooming, each plant was documented with high-resolution photography, including its dissected parts and the plant habit with relative scales. Photographs were mostly taken with Nikon cameras (D7100, D810) mounted on Manfrotto tripods, fitted with macro lenses (Micro Nikkor 60mm f/2.8, Micro Nikkor 105mm AF/ ED f/2.8, Sigma DG Macro HSM 105mm f/2.8). Floral details were captured with a Micro-Nikkor 60mm f.2.8 and Zeiss Luminar 63mm, 40mm, 25mm, and 15 mm, mounted on a Nikon PB6 bellows, or with a Leica Z16 APO Zoom microscope fitted with a Leica Planapo 1x Z-series lens. Images were post-edited for optimization using Adobe® Photoshop. Lankester Composite Digital Plates (LCDP) and other comparative plates of flowers were also prepared. Flowers and floral details were drawn using dissecting stereo microscopes Leica MZ9.5 and M80, fitted with Leica Planapo lenses and drawing tubes. Line drawings were mostly done using Rotring Rapidograph technical drawing pens with line widths of 0.1 or 0.2 mm on Fabriano, acid-free, smooth paper of 240g/m2. For some illustrations, the line drawing was prepared using an Apple® iPad and Procreate®, the electronic drawing successively printed on Fabriano paper with a HP Color Laser Jet Pro M452nw, and the printed drawing hand-dotted with a Rotring Rapidograph technical drawing pen.
Inflorescence terminology follows the proposal by Rojas-Alvarado et al. (2021). For all the descriptions, measurements were mostly taken under a dissecting stereoscope or with the aid of the electronic scale bars inserted in the high-definition images of the floral details. We specified the specimens on which each description is based. The vouchers were prepared by fixing entire or dissected flowers in FAA solution to be deposited at the JBL spirit collection. Additional Costa Rican material studied includes the revision of specimens deposited in other herbaria such as CR and USJ (acronyms cited follow Thiers 2023). Specific coordinates of cited specimens were omitted to prevent illegal poaching but can be provided upon request for scientific purposes.
Taxonomic treatment
Anathallis Barb.Rodr.
Anathallis funerea (Barb.Rodr.) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 115: 258. 2009. Basionym: Lepanthes funerea Barb.Rodr., Vellosia, ed. 2, 1: 118. 1891. Homotypic synonyms: Pleurothallis funerea (Barb.Rodr.) Cogn. in C.F.P.von Martius & auct. suc. (eds.), Fl. Bras. 3(4): 567. 1896. Specklinia funerea (Barb.Rodr.) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 95: 260. 2004. Panmorphia funerea (Barb. Rodr.) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 105: 156. 2006.
TYPE: [Brazil. Amazonas] Hab. os ramos delgados das arvores das mattas do Rio Yauapery, Floresce em Março [(holotype, lost; lectotype designated by Barros & Barberena, Rodriguésia 61(1): 128. 2010: fig. D, tab. 200, vol. 3 (J. Barbosa Rodrigues, Iconographie des Orchidées du Brésil, reproduced in Springer (1996, p.258)].
Heterotypic synonyms: Pleurothallis breviscapa C.Schweinf., Bot. Mus. Leafl. 3: 79. 1935. Anathallis breviscapa (C.Schweinf.) Pridgeon & M.W.Chase, Lindleyana 16(4): 248. 2001. Specklinia breviscapa (C.Schweinf.) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 95: 259. 2004. TYPE: British Guyana. June 1897, E.F. im Thurn 181 (holotype, K!).
Pleurothallis praemorsa Luer, Selbyana 2: 388. 1978. Specklinia praemorsa (Luer) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 95: 263. 2004. TYPE: Ecuador: Pastaza: epiphytic in rain forest about 20 km east of Puyo, alt. ca. 600 m, 3 Aug. 1977, C. Luer 1813, J. Luer & J. Brenner (holotype, SEL!).
Description: Based on D. Bogarín 10298.
Epiphytic herb, caespitose, small, to 5 cm tall. Roots flexuous, glabrous. Ramicauls erect, cylindrical, 0.6-2.0 cm long, 1 mm in diameter, with two internodes, the basal one shorter, covered by papery sheaths. Sheaths tubular, truncate to obtuse, ribbed, the basal one imbricate, the apical one shorter than the internode. Leaves elliptic to obovate, erect, 3.0-4.5 × 0.7-0.8 cm, base cuneate into a short petiole, apex acute, apiculate forming a minute tridentate apex. Inflorescence terminal at the apex of the ramicaul, with 1-4 successive multi-flowered coflorescences of 1.5-3.2 cm long, each one producing up to 4 successive brownish to greenish flowers. Pseudopeduncle 1-2 cm long, minutely pubescent, with a 3 mm long papery, acute, tubular bract at the base. Floral bracts 1.5-2.0 mm long, acute, minutely pubescent. Ovary subclavate, 0.75-1.0 mm long, minutely glandular, articled to a pedicel 2-5 mm long, minutely glandular. Dorsal sepal elliptic to ovate, acute, 12 × 4 mm, yellowish. Lateral sepals elliptic to ovate, acute, 11.0-12.0 × 3.0-3.5 mm. Petals yellowish with minute irregular brownish margins, acute, elliptic, 6.5-7.0 × 2.5 mm. Lip narrowly obovate, 4 × 1 mm, greenish, brownish to the margins, with a pair of auricles at the base, with short rounded and erected lateral lobes near the base, the middle lobe elliptic, with a truncate to emarginated apex. Column yellowish, 2 mm long, with a longhooded apex, formed by two laterals and one dorsal long acute tooth; stigma ventral. Anther cap ventral, yellowish, 0.7 mm wide; pollinarium composed of two pollinia, 0.3 mm wide, with a pair of caudicles. Fruit not seen.
Etymology: Presumably from the Latin fūnereus “funeral” in reference to the dark color of the flowers.
Distribution: Costa Rica, Panama, Guyana, Guyana Francesa, Venezuela, Ecuador, Peru, Bolivia, and Brazil.
Habitat in Costa Rica: Epiphytic in secondary wet forest on the Caribbean slope of the Cordillera Volcánica Central at about 750 m in elevation.
Phenology: Flowering in October and November under cultivation.
Costa Rican material studied: Limón: Pococí, Guápiles, Bellavista, ca. 3.8 km al sur de la Escuela La Guaria de Bellavista, 746 m, orillas de un afluente del Río Blanquito, bosque muy húmedo tropical, epífitas en bosque secundario, 8 jun. 2013, D. Bogarín 10298 (JBL-A0740 spirit!, Fig. 1).

Anathallis funerea is distinguished by the caespitose habit with minutely pubescent inflorescences as long as the leaf, producing single-successive brownish to greenish flowers, and the lip with a truncate apex. Anathallis angulosa (Luer & Hirtz) Luer is superficially similar, however it is distinguished from A. funerea by its glabrous inflorescence (vs. minutely pubescent), the acuminate petals (vs. acute), and the ciliate margins of the lip (vs. not ciliate?). The lip and the particular truncate apex of the lip in A. funerea are reminiscent of A. peroupavae (Hoehne & Brade) E.Barros, but, the latter is distinguished by the glabrous inflorescences and the pubescent petals (vs. minutely pubescent inflorescence and glabrous petals).
Dracula Luer
Dracula maduroiLuer, Monogr. Syst. Bot. Missouri Bot. Gard. 95: 234. 2004.
TYPE: Panama. Bocas del Toro: Velorio area, alt. 1100-1500 m, 10 Aug. 2003, A. Maduro & E. Olmos 410 (holotype: MO), C. Luer illustr. 20534.
Description: Based on N. Belfort-Oconitrillo 380 & K. Gil-Amaya, D. Bogarín 4405 et al., and K. Gil-Amaya 246.
Epiphytic herb, densely caespitose, up to 15 cm tall. Roots slender, ca. 1.2 mm in diameter. Ramicauls stout, erect, 1.30-3.20 cm long, enclosed by 2-3 tubular sheaths. Leaves erect to suberect, thinly coriaceous, carinate, narrowly linear-obovate, acute, 10.3-15.6 cm × 1.3-1.5 cm, gradually narrowed below into the conduplicate base. Inflorescence with successive multiflowered coflorescences, each of them distantly and successively few-flowered. Pseudopeduncle slender, slightly descending, 10-17 cm long; floral bract tubular, acuminate, green, 7-10 mm long; ovary verrucose, round in cross section, brown, 5-6 mm long, articled to a pedicel 11-15 mm long. Flower pendant, caudate, expanded, densely pubescent adaxially, sepals white with purple tails turning dull yellow towards the apex, slightly spotted purple brown near the base, suffused with purple-pink towards the basal portion of the tails, petals light yellow, turning bright yellow towards the verrucose apical portion, lip white, column white turning cream-yellow towards the apical portion. Dorsal sepal broadly ovate, shallowly concave from the middle to the basal portion below the column, 8.8 × 9.3 mm without the tail, connate to the lateral sepals for 4 mm to form an expanded flower, the apex obtuse, contracted into a slender, filamentous tail, 3.71-4.37 cm long. Lateral sepals 11.7-12.3 mm long without the tail, connate to ca. 11 mm to form a broad and bipartite synsepal 14.7 mm wide, shallowly concave near the base below the lip, the apices obtuse, contracted into slender tails 3-4 cm long. Petals cartilaginous, oblong, 3 mm long, l.5 mm wide, the apex obtuse, papillose, indistinctly bivalvate, the margin obtuse, denticulate, the outer lamina rounded, denticulate. Lip subpandurate, truncate, 4.7 × 2.2 mm, the hypochile oblong, 2.8-3.3 × 2.2 mm, hinged to the column-foot by a thin membrane, consisting of a deeply concave depression at the base, with erect, obtuse, marginal angles, cleft centrally; the epichile semicircular, 1.5-1.9 × 2.2 mm, obtuse, concave with slightly incurved margins becoming more evident when dehydrating, with 3 parallel, well-developed lamellae within, flanked by a few, vestigial, radiating carinae. Column arcuate, stout, semiterete, 3.7 mm long, with a stout foot ca. 2 mm long, at the end of the column, the clinandrium dorsally long hooded with a pair of small acute teeth. Anther cap cucullate, 2-celled, translucent white. Pollinarium composed of two pollinia, obovate, laterally flattened, ventrally depressed. Fruit not seen.
Etymology: Named for Andrés Maduro of Finca Dracula in the province of Chiriquí, Panama (Luer 2004).
Distribution: Known to occur in Costa Rica and Panama.
Habitat in Costa Rica: Epiphytic in secondary pluvial premontane and low montane forests along the Caribbean slope of the Cordillera Volcánica Central and northern foothills of the Cordillera de Talamanca, between 1150 and 1650 m.
Phenology: Blooming from July to February.
Costa Rican material studied: Alajuela: Sarchí, Bajos del Toro, Reserva Biológica Bosque de Paz, sendero Jaulares, 1540 m, bosque premontano en regeneración, creciendo como epífita sobre un árbol a una altura de ca. 3 m sobre el suelo, con una flor abierta, 27 jul. 2019, N. Belfort-Oconitrillo 380 & K. Gil-Amaya (JBL-A0520 spirit!, Fig. 2). Cartago: Paraíso, Orosi, Tapantí, Parque Nacional Tapantí, mar. 2021, K. Gil-Amaya 246 & G. Villalobos (JBL-B2286 spirit!). Heredia: Sarapiquí: Vara Blanca-Horquetas, camino al cráter del Cerro Cacho Negro, entre las riveras del Río Molejón y Río Cacho Negro, 1150 m, bosque pluvial montano bajo, epífitas en bosque primario a orillas del río, 10 abr. 2008, D. Bogarín 4405 et al. (JBL-D3523 spirit!).

The relatively small, densely pubescent white flowers and light-yellow petals distinguish this species among Dracula species present in Costa Rica. It can be compared to Dracula olmosii from Panama, but this species grows at a higher elevation, ca. 2000 meters, and exhibits slightly broader leaves and white petals. In comparison to its South American counterparts, it shares morphological similarities with the Colombian species D. posadarum Luer & R.Escobar and D. sergioi Luer & R.Escobar, but differs in having all-white, densely pubescent sepals with twice longer tails, and with the three lamellae of the epichile of the lip well-developed. Other Ecuadorian species with white sepals and purple sepaline tails include D. lotax (Luer) Luer, D. papillosa Luer & Dodson and D. rezekiana Luer & R.Hawley, but D. maduroi is distinguished by its expanded flowers, completely white sepals, short and dense pubescence, indistinctly bivalvate yellow petals, and the lip white with the epichile concave and the margins slightly incurved.
Dryadella Luer
Dryadella greenwoodianaSoto Arenas, Salazar & Solano, Icon. Orchid. (Mexico) 5-6: t. 550. 2002[2003].
TYPE: México. Oaxaca: Distrito de Ixtlán, km 46.7 del camino Ixtlán de Juárez-Talea de Castro, en la desviación a Tanetzé de Zaragoza y Juquila Quijanos, 2000 m, bosque de pino-encino-liquidámbar en filo de la loma; 25 mar. 2000, prensado de material cultivado, 2 jul. 2001, M. Soto et al. 9439 (holotype, AMO).
Description: Based on D. Bogarín 11108 et al. and A. P. Karremans 8870 & M. Contreras Fernández.
Plant epiphytic, caespitose or shortly rhizomatous, to 3 mm tall; rhizome to 2 mm long. Roots slender, to 1 mm in diameter. Ramicauls erect, to 3 mm long, enclosed by 2-4 thin, tubular sheaths to 3 mm long, the annulus to 0.3 mm. Leaves suberect, very thick (to 2.5 mm thick), coriaceous, conduplicate, elliptic, subacute, emarginate with a short apiculus, green, abaxially purple, 0.6-2.1 × 0.3-0.7 cm, narrowing into a subpetiolate base. Inflorescence with successive multi-flowered coflorescences, each of them with flowers opened at once. Pseudopeduncle short, to 2-4 mm long, rachis to 5 mm, floral bracts thin, 2.5-3.0 × 2.2. Ovary to 1.8 mm long, smooth, angulate, keeled, articled to a pedicel 2-4 mm long. Flowers the sepals greenish yellow with purple spots mostly along the veins, petals translucent greenish yellow with a red stain at the base and red spots towards the apex, the lip reddish. Dorsal sepal ovate, acute, concave, 4.5-4.7 × 2.6-2.8 mm, connate to the lateral sepals for 0.5 mm to form a sepaline cup. Lateral sepals ovate, acute to attenuate, thickened, connate for about 1.2 mm, 4.2-4.3 × 1.7-1.8 mm, with a transverse callus above the decurved base. Petals subpandurate to subhastate, acute with an obtuse angle on the upper margin and a subacute angle on the labellar margin, 1.7-1.8 × 1.2-1.3 mm. Lip unguiculate, arcuate, transversely ovate, hinged to the apex of the column-foot, 2.0-2.1 × 1.2-1.3 mm, rounded, decurved at the apex, the blade transversely obovate, auriculate, the disc with a pair of thick keels, the claw oblong, auriculate at the base, 8 mm long. Column subcylindrical, arcuate, erose at apex, to 3 mm long, with two acute, descending, stigmatic wings, with a conspicuous, canaliculate foot to 1.2 mm long, stigma ventral; anther subapical, anther cap cucullate, verrucose at the apex. Pollinarium composed of two pollinia, obovoid, with granulose caudicles. Fruit not seen.
Eponymy: Named after the Canadian Edward Warren Greenwood, one of the most influential contemporary orchidologists in Mexico.
Distribution: It has been recorded in Mexico, Guatemala, El Salvador, and Costa Rica.
Habitat in Costa Rica: Plants grow epiphytically in montane wet forest above 2400 m.
Phenology: Flowered in May and June in the wild and under cultivation.
Costa Rican material studied: Alajuela: Poás, San Juan, ca. 1 km antes de la entrada al Parque Nacional Volcán Poás, 2480 m, epífita en el tronco de un árbol aislado de Calyptranthes pittieri Standl. (Myrtaceae) en un potrero a orillas de la carretera, bosque pluvial montano, 19 may. 2014, D. Bogarín 11108, M. Fernández & L. Sandoval (JBL-illustration, Fig. 3-4A).


A. Dryadella greenwoodiana Soto Arenas, Salazar & R.Solano (D.Bogarín 11108). B. Dryadella guatemalensis (Schltr.) Luer (A.P.Karremans 3642).
Dryadella comprises six species in Costa Rica. Among the species in the country, D. greenwoodiana differs by the elliptic, very thick (to 2.5 mm thick), coriaceous leaves in contrast to the other five species that show narrowly linear or elliptic, obovate leaves less than 1.5 mm thick. The plants are also recognized by their adaxially green and abaxially purple leaves, usually overlapping to form compact, dense, somewhat repent plants (Soto Arenas et al. 2002). A BLAST search using molecular data from the nrITS available in GenBank (https://www.ncbi.nlm.nih.gov/genbank/) suggests that D. greenwoodiana is related to D. susanne (Pabst) Luer from Brazil, which shows thick leaves, and both form compact plants with overlapping leaves.
In Costa Rica, the habitat of D. greenwoodiana is very different from that of the other species of the genus. Dryadella greenwoodiana inhabits the montane forests at high elevations above 2400 m, while the other species inhabit premontane and forests below 650 m. Dryadella greenwoodiana has been observed above Sacramento de Barva, towards Volcán Barva in Heredia, and in the vicinity of Madreselva, El Guarco in Cartago. However, we were unable to prepare vouchers of these specimens
because they were infertile plants.
Isochilus R.Br.
Isochilus latibracteatus fo. albescens Karremans & Salguero, forma nova
TYPE: Cartago. Turrialba: Santa Teresita, Guayabo, Monumento Nacional Guayabo, alrededores de los senderos principales, 1128 m, bosque muy húmedo, epífitas, 30 sep. 2015, floreció en cultivo, preparada el 2 jul. 2020, A.P. Karremans 6720, N. Belfort-Oconitrillo & A. Morales (holotype, JBLJ1069 spirit!) (Fig. 5).

Diagnosis: A forma typica floribus albis versus lilacino-purpureos vel rubropurpureos recedit.
Description: Based on A .P. Karremans 6720.
Plant, epiphytic, cespitose, erect, up to 45 cm tall. Roots, dense, thick, fleshy, brownish, 3-5 mm in diameter. Rhizome, short, scandent. Stems, numerous, erect, 2-4 mm wide, covered by brownish leaf-sheaths, verrucose, especially the basal sheaths. Leaves, distichous, linear to linear-lanceolate, erect, decreasing in length as they approach the apex, apex conspicuously cleft, asymmetrically bilobed, 3.2 cm × 4.0 mm, articulated with their sheaths, inconspicuously carinate in the back, green. Inflorescence, terminal, raceme, dense, 4-5 cm long, 5-10 successive flowers. Floral bracts, imbricated, scarious, tubular in the base, oblanceolate, obtuse, 10-13 mm long, pale brown. Ovary, with a short pedicel, subtrigonous, ventrally canaliculated, pale green, completely covered by bracts, 5.0-6.0 mm × 2.5-3.0 mm including the pedicel. Flowers, white, with a magenta linear spot on the middle of the lip, 7-10 mm long, tubular-campanulate. Sepals, connate in the first basal quarter, 9.0-10.0 × 3.5-4.5 mm. Dorsal sepal, oblong, obtuse to rounded. Lateral sepals, gibbous at base, lanceolate to oblong-laceolate, prominently carinatewinged dorsally. Petals, elliptical-rhombic, reflexed apically, 8-9 × 3-4 mm. Lip, linear-oblanceolate, 9.0-10.0 × 2.5-3.0 mm, slightly constricted near the middle, apex obtused to rounded, with a sigmoid, saccate claw of 0.7-1.0 mm. Column up to 6 mm long, with a short inconspicuous foot, subclavate, ventrally canaliculate, clinandrium with oblong to ovate lateral wings 0.8-1.0 mm and an oblong-triangular tooth in the middle of 0.5 mm long, stigmatic cavity ventral, rostellum liner-triangular, acuminate. Anther cap, white, terminal, operculate, suborbicular, dorsiventrally compressed, 4-celled, cordate in the base, 1.5 × 1.5 mm. Pollinarium composed of four pollinia, in subequal pairs, obovate, elongate, and lateral compressed, pale yellow to whitish, 0.9-1.0 × 0.3-0.4 mm, attached to ovate-lanceolate hyaline caudicles with a keel. Fruit not seen.
Etymology: From the latin albēscēns “becoming white, whitish” in reference to the white color of the flowers, which contrasts with the typical form of the species.
Distribution: Known only from Costa Rica.
Habitat in Costa Rica: Very wet humid premontane tropical forest.
Phenology: Flowering in June and July.
Most species of Isochilus have either purpleor orange-colored flowers. The white flowers of Isochilus latibracteatus fo. albescens contrast notoriously with the violet-red or magenta found in the typical form of this species. Only three species of Isochilus have been formally reported to occur in Costa Rica: I. linearis (Jacq.) R.Br., I. carnosiflorus Lindl. and I. chiriquensis Schltr. (Dressler 2003). The reason why we want to describe formally this new form is because it’s the only one known in Costa Rica, the typical form of Isochilus lactibracteatus is not known to occur yet in the country. Also, it is known that color polymorphism in orchids can have different ecological and evolutionary implications (Dormont et al. 2019).
The species is easily distinguished from I. linearis by the dark green leaves that are shorter toward the apex and, most important, the wide bracts that cover completely the pedicellate ovary. This new form has a very attractive white perianth and a magenta linear spot in the middle of the lip.
Lepanthes Sw.
Lepanthes acarinaLuer, Phytologia 54(5): 326. 1983. TYPE: Ecuador. Pichincha: epiphytic in cloud forest near Río Silante, Finca Canchacato, alt. ca. 2000 m, 28 oct. 1979, C. Luer, J. Luer & A. Hirtz 4399 (holotype, SEL).
Description: Based on D. Bogarín 12501 & K. Gil-Amaya and K. Gil-Amaya 202.
Plant minute, epiphytic, caespitose, erect herb, up to 2.3 cm tall. Roots filiform, flexuous, slender, 0.2 mm in diameter. Ramicauls slender, erect, 8-14 mm long, enclosed by 3-6, tightly fitting, scabrous, lepanthiform sheaths. Leaves coriaceous, elliptical to ovate, obtuse, conduplicate, apiculate, abaxially with purplish veins, 7.0-11.7 × 4.6-6.9 mm, the rounded base contracted into a petiole ca. 1 mm long, the apex obtusely shortly cuspidate, excise, with the tip of the central vein protruding abaxially within the sinus. Inflorescence developed beneath the leaf with successive multi-flowered coflorescences, 3.6-8.5 mm long, each of them distichous, successively flowered. Pseudopeduncle up to 6.3-8.0 mm long, as long or longer than the leaf. Floral bracts triangular-ovate, muriculate, yellowish brown, 0.8-1.0 mm long. Pedicels 1.0-1.5 mm long, persistent. Ovary to 1 mm long, with prominent, slightly spiculate ribs. Flowers with yellowish-white, translucent sepals, the apex of the dorsal sepal tinged with purple, petals and lip purplish pinkish. Dorsal sepal broadly ovate-triangular, subacute, serrulate-ciliate and dorsally carinate, connate to the lateral sepals for about 0.6 mm, 2.0-2.1 × 1.3-1.5 mm, 3-veined. Lateral sepals ovate, acute, serrulate, dorsally carinate-denticulate, connate for 0.8 mm, 1.7-1.9 ×1.0-1.1 mm, 2-veined. Petals bilobed, transversely oblong, minutely ciliate, 1.1-1.2 × 0.3-0.4 mm, upper lobes oblong, oblique, subtruncated, lower lobes narrowly oblong, obtuse, smaller than the upper lobes. Lip bilaminate, rounded, adnate to the column, the blades ovate, lunate, microscopically pubescent, 0.5-0.7 × 1.0-1.1 mm, connectives cuneate, up to 0.5 mm long; body narrow, connate to the base of the lip, appendix conspicuous for the genus, constricted above the middle, elliptic to oblong, the base concave, and the apex deflexed, pubescent. Column stout, terete, 1 mm long, with the anther dorsal, the stigma ventral. Pollinarium composed of two pollinia, ovoid, yellow. Anther cap cucullate, 1-celled. Fruit not seen.
Etymology: Named after the order of the mites Acarina which is derived from the Greek ακαρής, akarés, “tiny”, in reference to the little, red, prickly flowers (Luer 1983).
Distribution: Costa Rica, Colombia, Ecuador, Peru, and Bolivia.
Habitat in Costa Rica: Epiphytic in humid lower montane forest on Quercus spp. (Fagaceae) and Cupressus lusitanica Mill. (Cupressaceae), between 1800-1900 m.
Phenology: Flowering between July and October.
Costa Rican material studied: Cartago: Quebradilla, cruce a Alto Mata de Caña, carretera entre Tablón y Copalchí, 1891 m, bosque húmedo montano bajo, epífita en bosque secundario, 11 oct. 2019, D. Bogarín 12501 & K. Gil-Amaya (JBL-illustration!; Fig. 6); same locality, 7 jul. 2019, K. Gil-Amaya 202 (JBLA0384 spirit!).

This species is widespread in the tropical Andes and is documented here for the first time in Southern Central America. Lepanthes acarina is the third species of the genus known to range from Costa Rica and Panama to Colombia and Ecuador (Bogarín et al. 2016). It is easily distinguished by its diminutive habit (plants <3 cm tall), with inflorescences developed beneath the leaves, often longer than or as long as the leaves themselves. The flowers are minute, yellowish-white with purplish-pinkish petals and lip. The sepals are ciliate, with the dorsal sepal tinged with purple. The lunate lip blades embrace the column, and the appendix is conspicuous, constricted above the middle, with a concave base and a pubescent apex that is deflexed.
Within the populations, we observed cleistogamous plants, with some flowers opening occasionally. While L. acarina shares a similar plant habit with the widespread L. eximia Ames, which is often cleistogamous, they can be differentiated by the linear petals (as opposed to transversely bilobed, oblong petals in L. acarina) and the subquadrate, widely lobed lip with lobes extending far from the column (versus ovate, short lip blades in L. eximia). Plants of L. acarina typically grows on mossy twigs alongside other pleurothallids such as Dracula erythrochaete (Rchb.f.) Luer and Stelis spp.
Lepanthes pexaLuer, Lindleyana 2(4): 181. 1987.
TYPE: Panama. Prov. of Bocas del Toro: epiphytic in cloud forest between Fortuna and Chiriquí Grande, alt. 1000 m, 17 Feb. 1985, C. Luer et al. 10622 (holotype, MO-5122325!, barcode 150103).
Description: Based on D. Bogarín 8243.
Plant epiphytic, caespitose, erect, up to 16 cm tall. Roots slender, flexuous, up to 2.3 cm long, 1 mm in diameter. Ramicauls slender, erect to suberect, 6.0-11.5 cm long, enclosed by 5-7 pale green to brownish with age, closely adpressed lepanthiform sheaths, the ostia microscopically ciliate, ovate, acute to obtuse. Leaves elliptical, emarginate, apiculate, coriaceous, papillose, green, lustrous, 4.1-5.7 × 2.1-2.4 cm, the obtuse base narrowing into a petiole ca. 5 mm long. Inflorescence born beneath the leaf with successive, distichous multi-flowered coflorescences, up to 3.3 cm long, each of them shorter than the leaves, with successive flowers. Pseudopeduncle up to 2.4 cm long, rachis up to 8 mm long; floral bracts pale green to brownish with age, ovate, acute, muricate, pilose, 1.5-2.0 mm long; pedicels 2-3 mm long, persistent; ovary microscopically papillose, 1.0-1.5 mm long. Flowers with yellowish-green, translucent sepals, the petals light orange, basally vinaceous, the upper lobe deeply bordered with wine color, the lower lobe lightly vinaceous, the lip lilac, the column light orange to whitish. Dorsal sepal ovate, acute, abaxially 1-carinate, entire, connate to the lateral sepals for about 0.5 mm, 3.0 × 2.1 mm, 3-nerved. Lateral sepals obliquely ovate, acute, apically divergent, abaxially 1-carinate, connate for 1.1 mm, 2.8 × 1.4 mm, 2-nerved. Petals transversely bilobed, microscopically papillose, 1.3 × 4.0 mm, the upper lobe obliquely ovate, obtuse, converging, 2.2 mm long, the lower lobe narrow, obliquely ovate, obtuse, subcarinate along the vein, diverging at the apex, 1.8 mm long. Lip bilaminate, adnate to the column base, 1.5 × 2.0 mm, not exceeding the column length, the blades elliptic, obtuse, apically ciliate; the connective oblong, 0.5 × 1.0 mm; the body oblong, thick, adnate to the column base, 0.4 × 0.2 mm; the appendix cylindric, obtuse, pubescent, 0.15 mm long. Column cylindric, 1.3 mm long, exceeding the length of the lip; the anther apical; the stigma ventral. Pollinarium consisting of two pollinia, claviform, 0.55 × 0.28 mm, joined at the base by a rounded viscidium. Anther cap, subrounded, abaxially concave, 0.15 × 10 mm. Fruit not seen.
Etymology: From the Latin pexus, “combed”, referring to the appearance of the densely distichous pedicels on the rachis (Luer 1987).
Distribution: The species is known from Costa Rica and Panama.
Habitat in Costa Rica: Plants were found growing as epiphytes on the trunks of Cupressus lusitanica (Cupressaceae) at the edge of the lower montane pluvial forest on the Pacific watershed of the Central Valley at 1484 m of elevation.
Phenology: Flowered in February and November under cultivation.
Costa Rican material studied: San José: Moravia, San Jerónimo, 1484 m, calle Carrillo y Tornillal, floreció en cultivo en el Jardín Botánico Lankester, preparado 18 Nov. 2010, D. Bogarín 8243 &. D. Matamoros (JBL-D4610 spirit!, Fig. 7-8).


Lepanthes pexa is an erect herb with leaves shorter than the compact raceme. The petals have obliquely ovate lobes, while the lip, bilaminate in structure, does not exceed the length of the column. The lip consists of rose, elliptic blades with ciliate apices, an oblong body, a cylindric and obtuse appendix, and a short, cylindric column. The type specimen of L. pexa exhibits some differences in the petals compared to available Costa Rican material. For instance, its petals are acuminate at both proximal and distal ends (versus obtuse), while the labellum bears cilia at the apex of the blades (versus non-ciliate). Nevertheless, plants from Costa Rica share the characteristic rose lip, which is unusual within the genus in Costa Rica and Panama. We consider these differences variable characters until more material is available to assess morphological variation within populations.
According to Luer (1987), L. pexa resembles Lepanthes turialvae Rchb.f., a species found in Costa Rica and Panama. However, L. pexa can be distinguished by the carinate lower lobe of the petals (versus noncarinate) along the vein and the converging upper lobe (versus divergent).
Masdevallia Ruiz & Pav.
Masdevallia smallmaniana Luer, Monogr. Syst. Bot. Missouri. Bot. Gard. 87: 505a. 2002. Luzama smallmaniana (Luer) Luer, Monogr. Syst. Bot. Missouri. Bot. Gard. 105: 10. 2006.
TYPE: Ecuador[?]. without locality, alt. 2000-2500 m, obtained from New World Orchids by the British National Collection of Masdevallia, flowered in cultivation at Royden Orchids in Grand Missenden, England, Feb. 2002, R. Barrow RO11 (holotype, barcode MO-277324!).
Heterotypic synonyms: Masdevallia driessenianaLuer & Sijm, Harvard Pap. Bot. 23: 48. 2018. TYPE. Panama. Chiriquí: near Amistad, 1300 m, collected by P. Dubbeldam and A. Sijm, 2003, flowered in cultivation by Wiel Driessen in Panningen, The Netherlands, February 2013, A.P. Sijm 20130208 (holotype: MO). Masdevallia rostriflora Luer & Sijm, Harvard Pap. Bot. 23(1): 50. 2018. TYPE: Panama. Chiriquí: near Amistad, 1800 m, collected by P. Dubbeldam and A. Sijm, 2003, flowered in cultivation by Wiel Driessen in Panningen, The Netherlands, Feb. 2013, A.P. Sijm 20130204 (holotype: MO).
Description: Based on Bogarín 10767 et al.
Plant up to 7.5 cm tall, epiphytic, caespitose; thick roots. Ramicauls slender, erect, 5.4-11 mm long, enclosed by one thin, tubular sheath papyraceous, 5-10 mm long. Leaves light green darkening towards the apex, erect, coriaceous, cuneate, oblanceolate, obtuse, 54-65 × 8.5-11.0 mm including a gradually narrowed indistinct petiole ca. 8-15 mm long. Inflorescence with semierect successive single-flowered coflorescences, up to 5 cm long. Pseudopeduncle light green, with light wine color spots from the base to the halfway, 2.7-4.0 cm long, with a tubular bract near the base; floral bract light green, tubular, 1 cm long; ovary light green, 5 × 2 mm wide, articled to a 6 mm long pedicel; sepals greenish white, lightly suffused with red-brown at the base, glabrous. Dorsal sepal oblong, 20 × 4 mm including the thick tail 9 mm long, connate to the lateral sepals for 6 mm into a cylindrical tube, the free portion triangular, acute, attenuate into an erect, light olive-green tail, more pigmented towards the apex. Lateral sepals oblong, oblique, acute, 20 × 8 mm, connate 8 mm, synsepal expanded, the free portions narrowly triangular, acute, with light olive green, more pigmented towards the apex, fleshy, decurved tails ca. 12 mm long. Petals light olive green, red-brown above the middle, obovate below the middle, contracted into a acuminate-subulate apex suffused with red-brown spots, 13 × 3 mm. Lip greenish, red-brown above the middle, protruding, 15 × 3 mm, oblong below the middle, contracted above the middle into narrowly linear-subulate apex, with a midline callus extending forward onto the apex. Column light green, semiterete, 7 mm long. Anther greenish white cucullate; Pollinarium composed of two pollinia. Fruit not seen.
Etymology: Named for Don Smallman of Halpringham, Norfolk, England, who cultivates this species in the British National Collection of Masdevallia.
Distribution: Costa Rica, Panama, and doubtfully in Ecuador.
Habitat in Costa Rica: Epiphytic in low montane rainforest and in isolated pasture trees on the Cordillera de Talamanca at 2162 m.
Phenology: Flowering in May and December.
Costa Rican material studied: Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 15 km al noreste de Lucha, Sitio Tablas, en potreros de la Finca SandíHartmann “El Capricho”, 2162 m, epífitas en árboles aislados en bosque pluvial montano bajo, 11 dic. 2013, D. Bogarín 10767 et al. (JBL-D6293 spirit!; Fig. 9).
In Costa Rica, certain populations of Masdevallia cupularis Rchb.f. exhibit individuals that bear flowers that never reach anthesis and are self-pollinated while still in bud. This phenomenon is often accompanied by the development of peloric buds and flowers, wherein the floral segments display aberrant morphology. Pelorism typically denotes an abnormality where flowers appear to revert to an earlier, ancestral form. The characteristic bilateral symmetry of orchid flowers is often lost in peloric flowers, resulting in actinomorphic individuals (Mondragón-Palomino & Theißen, 2009). Taxonomists have identified three such aberrant individuals as distinct species, named M. smallmaniana Luer, M. driesseniana Luer & Sijm, and M. rostriflora Luer & Sijm (Karremans et al. 2019; Karremans 2023). All three are peloric or pseudopeloric, representing individuals of a single species. Oses (2017) found individuals of M. cupularis and M. smallmaniana as sisters within the same clade. Whether the peloric individuals form a stable population and can be considered a distinct species from M. cupularis needs to be assessed through population genetics. Meanwhile, M. smallmaniana is formally recorded in the flora of Costa Rica to acknowledge the presence of these aberrant populations.
Masdevallia striatella fo. pallens Belfort, forma nova
TYPE: Costa Rica. Alajuela: Sarchí, Bajos del Toro, Reserva Biológica Bosque de Paz, creciendo en el Jardín de Orquídeas Dr. Stephen Kirby, 1534 m, 29 oct. 2023, N. Belfort-Oconitrillo 938 (holotype, JBL-spirit; LCDP voucher, Fig. 10).

Diagnosis: A forma typica floribus albescentibus sepalibus venis rosaceis pallentis notatis, petalis labelloque plerumque albescentis (versus venas sepalorum atreopurpureas et labellum atropurpureum) recedit.
Epiphytic, caespitose, herb, 15-20 cm tall. Ramicauls green, terete, slender, erect to suberect, 5-10 cm long. Leaves obovate and subpetiolate, 6.7-14.1 × 0.7-1.0 cm. Inflorescence with successive multiflowered coflorescences, 7-13 cm long, producing up to two flowers. Pseudopeduncle 6-8 cm long, erect. Pedicels 7-10 mm long. Ovary cylindrical, 5 mm long, longitudinally channeled, with evident black dots. Floral bract triangular-ovate, brownish, membranaceous, acute, tubular, apically loose, involving most of the pedicel, 5-7 mm long. Flowers with sepals connate, tubular, white with nine veins light pink (three in each sepal), the sepaline tails green, the petals white with the middle vein pink, the lip pale with the base and the apex yellow, white column with light pink margins, white anther cap and yellow pollinarium. Sepals lanceolate, 2.01-2.18 × 0.57-0.62 cm, connate at the base for 0.7-0.8 cm, forming a tube 0.7-1.0 cm in diameter that opens towards the ends into three short sepaline tails of 5.5-6.0 mm long. Petals elliptical, narrow at the base, 6.5 × 2.5 mm, with an angle in the middle of the lower margin and with the apex mucronate, disposed on each side of the column, forming a narrow tunnel. Lip oblong, arched, 7.5 × 3.5 mm, base truncated, with two basal concave lobes and exudate as droplets in the central portion, with a minutely warty callus at the apex, bent slightly backward. Column elongated, straight, 6.5-6.7 mm long, shorter than the lip, with a foot of 3.5 mm long and a hook with a bifurcate apex, to which a thin layer of cells articulates the lip. Anther cap cucullate, white. Pollinarium composed of two pollinia, obovate, with a pair of whale-tail-shaped caudicles. Fruit not seen.
Etymology: From the Latin pallēns, meaning pale, in reference to the pale color of the flowers, especially due to the much lighter petals and lip.
Distribution: Known only from Costa Rica.
Habitat in Costa Rica: Premontane tropical forest in the Caribbean slope of the Cordillera Volcánica Central.
Phenology: Blooming from September to January, with a peak in November.
Masdevallia striatella Rchb.f. is known to occur in Costa Rica and Panama, representing one of the most common species of the genus and with the broadest geographical and ecological distribution (Luer 2006, Oses 2017). The typical form consists of whitish flowers with evident dark purple veins in the sepals, green or yellow-colored sepaline tails, and a dark purple lip with yellow in the base and the apex (Fig. 11). Although in the new form, the sepals still have purple veins but are much lighter, the most evident differences are the green tails, the white lip and the white petals with much pale, inconspicuous veins (Fig. 10C), contrasting with the dark purple lip and dark veins of the typical form (Fig. 11B). This new form became evident at Bosque de Paz Biological Reserve, where the pale form blooms synchronously with plants exhibiting the typical characteritics of the species.

Typical floral form of Masdevallia striatella Rchb.f. from Bosque de Paz Biological Reserve.
Maxillaria Ruiz & Pav.
Maxillaria hennisiana Schltr., Orchis 6(7): 117-118. 1912.
TYPE: Colombia. Heimat wahrscheinlich Kolumbien, von H. Hennis in Hildesheim importiert, blühte in der Sammlung des Herrn Baron v. Fürstenberg im Juni 1911, Herrn Baron von Fürstenberg s.n. (B, destroyed); lectotype, hic designatus, the illustration in the protologue (Schlechter 1912): tab. 26, fig. 10-17. Fig. 12.
Heterotypic synonym: Maxillaria shepheardii Rolfe, Bull. Misc. Inform. Kew 83. 1917. TYPE: Colombia. Chocó: Río Condoto, S. Shepeard s.n. (holotype, K).
Description: Based on L. Álvarez et al. 1229, L. Álvarez 499 & M. L. Morales, and L. Álvarez 335.

Original illustration of Maxillaria hennisiana Schltr. From Schlechter 1917: tab. 26, fig. 10-17.
An epiphytic, caespitose, pseudobulbous herb to 15 cm tall. Roots slender, flexuous, glabrous, ca. 1 mm in diameter. Rhizome with short internodes, covered by brown, nervous, papyraceous sheaths. Pseudobulbs transversely sub rectangular, thickly complanate, apically truncate, to 2.3 × 2.8 cm, monophyllous, covered at the base by 2-3 triangular, narrow, acute, glumaceous sheaths to 3 cm long, becoming dry-papyraceous with age. Leaves narrowly elliptic to oblong-elliptic, coriaceous, grass-green, acute to subobtuse, asimetrically bilobulate, distinctly keeled beneath, matte on both surfaces, 6.0-13.0 × 1.8-2.3 cm, narrowed at the base into a long, conduplicate-terete petiole to 3.5 cm long, ca. 0.5 cm wide. Inflorescences up to 8 per pseudobulb, each a single-flowered, erect raceme shorter than the leaves, 5-7 cm long, completely covered by 3-4 amplectent, tubular, apiculate, papyraceous, brown bracts to 2.8 × 0.8 cm. Floral bract glumaceous, brownish green, amplectant, obovate, acute, apically loose, strongly ancipitous, to 3 × 1 cm, completely covering the pedicel and ovary. Pedicellate ovary terete, rugulose, 3.0-3.2 cm long including the pedicel. Flowers with the sepals spreading, the petals porrect, the sepals yellow flushed brown toward the apex, the petal white, the lip yellow with orange midlobe, the column white, the anther reddish with dark purple crest. Dorsal sepal linear-ligulate, obtuse, carinate, the distal margins gently reflexed, 2.3-2.5 × 0.5-0.6 cm. Lateral sepals narrowly linear-ligulate, obtuse, shortly apiculate, weakly carinate, 2.4-2.6 × 0.5-0.6 cm, the bases forming a short mentum with the column foot. Petals obliquely narrowly lanceolate-subfalcate, subacuminate, porrect, the apex facing down, 1.7-2.0 × 0.4 cm. Lip 3-lobed, elliptic from a cuneate base, adnate to the base of the column foot, 1.2 × 0.6 cm, the disc provided with a ligulate, low callus ca. 5 mm long, extending almost to the insertion of the middle lobe; lateral lobes rounded, erect to flank the lower portion of the column; midlobe ellipticsubrounded, thickened, the margins crenulate. Column semiterete, thick, 1 cm long, dilated around the stigma, extending at the base into a narrow, cuneate foot ca. 4 mm long; the clinandrium cucullate, lacerate; the stigma ventral, broadly subquadrate, flanked by two large, converging, membranous flaps; the anther incumbent. Anther cap deeply cucullate, ovate, obtuse, provided with a distinct, narrow crest on the upper part, 2-celled. Pollinarium consisting of four pollinia, ovoid-complanate, in two pairs of different size, dorsiventrally superposed, on a horseshoe shaped stipe, scarcely distinct from the subtended, brownish visicidium. Fruit not seen.
Etymology: Dedicated to Wilhelm Hennis (1856- 1943), orchid grower in Hildesheim, who imported from Colombia the type plant later flowered in the collection of Baron Max (Maximilian) von Fürstenberg (1866-1925) in June 1911.
Distribution: Costa Rica to Ecuador.
Habitat in Costa Rica: The species has been observed so far exclusively in lowland, wet tropical forest in the Pacific region of Península de Osa, at elevation around 400 m, as an epiphyte in both primary and secondary vegetation.
Phenology: Flowering in February-March and in October-November.
Costa Rican material examined: Puntarenas: Golfito, Golfito, Parque Nacional Piedras Blancas, Fila Monge, 420 m. Bosque primario, con especies arbóreas predominantes como Peltogyne purpurea, Pentagonia osaensis, Magnolia wetteri, Virola nobilis, Chaunochiton kappleri, Tachigali versicolor, 17 mar. 2021, L. Álvarez 499 & M. L. Morales (JBL-B2626 spirit!) (Fig. 13). Golfito, Golfito, Parque Nacional Piedras Blancas, Fila San Josecito, 400 m, bosque muy húmedo tropical, bosque primario, 10 may. 2020, florecida en cultivo y preparada en el Jardín Botánico Lankester, 18 feb. 2022, L. Álvarez 335 (JBL-A0616 spirit!) (Fig. 14A). Golfito, Golfito, Cerros de Golfito entrando por la Gamba, propiedad de Conservación Osa, 400 m, bosque secundario, 7 jun. 2023, flowered in cultivation and prepared at Jardín Botánico Lankester, 28 oct. 2023, L. Álvarez 1229, M. J. Mata & F. Rodríguez (JBL) (Fig. 14B).


Maxillaria hennisiana belongs to a species complex centered around the South American M. porrecta Lindl., the first species surely referable to this the group, which was described in 1838 from Rio de Janeiro (Lindley 1838). This complex includes other species such as M. acostae Schltr., M. amparoana Schltr., M. angustisegmenta Ames & C.Schweinf., M. brunnea Rchb.f., M. colemanii Carnevali & Fritz, M. cryptobulbon Carnevali & J.T.Atwood, M. longiloba (Ames & C.Schweinf.) J.T.Atwood and M. ringens Rchb.f., among others. Christenson (2002) treated this complex as the section Amazonicae, excluding however M. hennissiana as a representative species. Morphologically, species whitin this group share characteristics with Maxillaria sect. Arachnites Christenson - distinguished primarily by the larger size of plants and flowers - and together they represent one of the largest and less studied groups in Maxillaria. In their classification of the subtribe Maxillariinae based on ITS molecular data, Szlachetko et al. (2012) recovered M. hennisiana within Maxillaria sensu stricto, forming a small clade with M. longiloba, M. angustisegmenta, M. cryptobulbon, and M. brunnea as the closest relatives. However, the phylogenetic analyses of Maxillaria s.s. by the authors revealed several polytomies and did not reflect recent attempts at sectional delimitations as those posthumously proposed by Christenson et al. (2013). Due to the difficulty in distinguishing between species of these groups, given their overall floral similarity and colors, it is unsurprising that M. hennisiana has been recorded as ranging from southern Mexico to Peru and Trinidad in the West Indies. According to the most recent reports, the species is likely restricted to Panama (Bogarín et al. 2015), Colombia, and Ecuador (Bernal et al. 2019). This is the first documented record of the species for the flora of Costa Rica.
Among similar species, M. hennisiana may be recognized by the compact plants with distinctly petiolate leaves and the peculiar shape of the pseudobulbs, which are transversely rectangular, thickly lenticular, and distinctly truncate apically (Fig. 15).

Maxillariella M.A.Blanco & Carnevali
Maxillariella guareimensis (Rchb.f.) M.A.Blanco & Carnevali. Lankesteriana 7(3): 528. 2007.
TYPE: Venezuela. Wagener s.n. (holotype, W-R?; isotype, G-00355233!). Syn.: Maxillaria guareimensis Rchb.f., Bonplandia 2: 16. 1854.
Description: Based on Karremans 6671 et al. and Karremans 7285 et al.
An epiphytic, cane-bearing herb to 1 m tall. Roots slender, flexuous, glabrous, ca. 1 mm in diameter. Rhizome with short internodes, covered by brown, papyraceous sheaths, up to 5 cm long between stems. Stems elongate, covered by foliaceous and papyraceous sheaths, 4.1-6.8 × 1.7-2.0 cm. Pseudobulbs not seen. Leaves narrowly elliptic, coriaceous, acute, asymmetrically bilobulate, distinctly keeled beneath, the blade 10.0-20.0 × 2.1-3.2 cm. Inflorescences from the base of the foliaceous sheath, several simultaneously appearing per stem, erect. Floral bract glumaceous, brown, amplectent, acute, apically loose, ancipitous, 1.0-1.5 cm, not covering the pedicel and ovary. Pedicellate ovary terete, rugulose, 2.5 cm long including the pedicel. Flowers concolorous yellow. Dorsal sepal elliptic, acute, shortly apiculate, 1.8-1.9 × 0.8 cm. Lateral sepals narrowly ovate, oblique, acute, shortly apiculate, apex revolute, 1.6-1.7 × 0.8 cm, the base forming a short mentum with the column foot. Petals narrowly ovate, acute, apex revolute, 1.5-1.6 × 0.8 cm. Lip linear-pandurate, hinged to the base of the column foot, 1.2 × 0.5 cm, the disc provided with a ligulate, low callus ca. 6 mm long, extending from the base to the center of the lip, apex truncate, slightly emarginate. Column semiterete, thick, 0.8 cm long, clavate, extending at the base into a narrow, cuneate foot ca. 3.5 mm long; the clinandrium cucullate, glandular; the stigma ventral, broadly subquadrate, flanked by two large, converging, membranous flaps; the anther incumbent. Anther cap deeply cucullate, ovate, obtuse, 2-celled. Pollinarium composed of four pollinia, ovoid-complanate, in two pairs of different size, dorsiventrally superposed, on a horseshoe shaped stipe, scarcely distinct from the subtended, brownish viscidium. Fruit not seen.
Etymology: The name presumably honors mount Guareima, close to Caracas in Venezuela.
Distribution: Trinidad and Tobago, Costa Rica, Venezuela, Colombia, Ecuador, Peru, Bolivia, Brazil.
Habitat in Costa Rica: Very wet premontane tropical forest.
Phenology: Blooming at least in June.
Costa Rican material examined: Cartago: Turrialba, Santa Teresita, Guayabo, Monumento Nacional Guayabo, alrededores de los senderos principales, 1128 m, bosque muy húmedo, epífitas, 20 jun. 2015, A.P. Karremans 6671 et al. (JBL-J0622 spirit!, Fig. 16). Turrialba, Santa Teresita, Guayabo, Monumento Nacional Guayabo, potreros al lado de la calle, a orillas del Río Lajas, 1.5 km Norte de la entrada principal, 1270 m, bosque muy húmedo, epífitas, 5 jun. 2016, A.P. Karremans 7285, G. Rojas-Alvarado & Orquideología UCR2016 (CR, USJ).

This seems to be a rather poorly known species despite its broad distribution. In Costa Rica, this rather voluminous orchid had somehow eluded decades of botanical exploration until recently discovered growing at the Monumento Nacional Guayabo, in Turrialba. Maxillariella guareimensis forms large masses, composed of multiple elongate stems, high on the thick tree trunks along the Lajas river that crosses the small natural reserve. Its cupped, yellow flowers are reminiscent of Maxillariella diuturna (Ames & C.Schweinf.) M.A.Blanco & Carnevali, a species with a significantly smaller plant stature and floral dimensions which occurs in the same area. Besides its large size, the newly recorded species can also be distinguished by the lack of pseudobulbs along the stem, and the neatly ordered, dense set of leaves.
Mormolyca Fenzl
Mormolyca gracilipes (Schltr.) Garay & M.Wirth, Canad. J. Bot. 37(3): 482. 1959. Bas. Cyrtoglottis gracilipes Schltr., Repert. Spec. Nov. Regni Veg. 7: 182. 1920.
TYPE: Colombia. Cauca: c. 1200 - Madero s.n. [holotype, B, destroyed; lectotype (hic designatus), Schlechter’s floral analysis published by Mansfeld (1929: Pl. 65, No.250)]. Colombia. Cauca: c. 1000 - Lehmann, F. C. 813 [neotype, AMES (Garay & M.Wirth 1959), here rejected and selected as epitype]. Note: We follow Pupulin et al. (2022) in recognizing Schlechter’s floral analysis reproduced by Mansfeld as original material and therefore reject Garay & M. Wirth neotype in favor of a lectotype. Article 9.12 of the Shenzhen code (Turland et al. 2018).
Description: Based on Bogarín 13092 & A. Acuña.
Plant epiphytic, cespitose, erect, up to 27 cm tall. Roots scatered, thick, fleshy, whitish, 0.5-1.5 mm wide. Rhizome short, creeping. Pseudobulbs clustered, oblong-ovoid, strongly complanate, unifoliate at the apex, with distichous, evanescent sheaths at the base, 2.5-5.5 × 0.8-1.3 cm. Leaves green, elliptic-oblong, acute, 5.5-11.0 × 1.0-1.5 cm. Inflorescences two or more, from the base of the pseudobulb, 1-flowered, elongate surpassing the leaf, erect, slender, peduncle about 3.0-6.0 cm long, with 3-4, brown, tubular sheaths. Flowers large for the plant, orange yellow. Dorsal sepal lanceolate-ovate, longacuminate with the upper margin involute, concave, 2.1-2.8 × 0.8-1.3 cm. Lateral sepals lanceolate, long acuminate, with the upper margin involute, larger than the dorsal sepal, 2.4-3.5 × 0.8-1.0 cm. Petals much shorter than the sepals, linear, sharply acute, vertically disposed, touching each other, 1.2-1.4 × 0.1-0.15 cm. Lip short, strongly convex in natural position, rhombic-obovate when expanded, with an indistinct indentation on each side, about the middle, abruptly acute, hairy at the tip, about 9.0-1.1 × 0.5-0.6 cm, with the central part very thickened. Column green, with brown-purple spots ventrally and at the base, small, slender, arcuate, clavate, footless, up to 0.7-1.0 cm long. Anther cap, orange, terminal, operculate, 2-celled, cordate at the base 1.6-1.8 × 1.2-1.4 mm. Pollinarium composed of four pollinia, in two subequal pairs, obovate, laterally compressed, yellow, 0.5-0.7 × 0.3-0.4 mm, attached to a wide horseshoe-shaped viscidium. Fruit not seen.
Etymology: From the Latin pedis “foot” and the Latin gracilis “slender, thin, slim”, probably alluding to the slender shape of the column.
Distribution: Costa Rica, Venezuela, Colombia, Ecuador, Perú, and Bolivia.
Habitat in Costa Rica: Very humid premontane tropical forest, in moderately warm climates.
Phenology: Flowering in November.
Costa Rican material studied: Limón: Siquirres, La Alegría, San Bosco, Calle La Iberia, cerca de orillas del río Destierro, límite entre La Alegría y Pocora de Guácimo, 350 m, bosque muy húmedo tropical transición a premontano, epífitas, 22 ago. 2020, D. Bogarín 13092 & A. Acuña, floreció en cultivo en JBL, 26 nov. 2020 (JBL-K0121!, JBL-L0029!; LCDP voucher, Fig. 17).

Mormolyca gracilipes was initially described as Cyrtoglottis gracilipes by Schlechter in 1920, based on a Colombian specimen to describe a new genus. Schlechter recognized the similarity with the genus Mormolyca at that time. However, he considered that the shape of the flower with its long and pointed floral parts, in addition to its geographical disjunction, showed that it was a different genus (Garay & Wirth 1959). The geographical separation between both was lost with the discovery of M. peruviana C.Schweinf. (Schweinfurth 1944), which extended the range of the genus Mormolyca from Central America to South America. For this reason and considering a new species with intermediate characteristics, Mormolyca polyphylla Garay & Wirth (1959), concluded that there were not enough reasons to separate the genera. Thus, becoming synonyms.
Mormolyca gracilipes could be easily confused at first sight with M. schweinfurthiana Garay & Wirth, however, it is distinguished by its simple (vs. trilobed), fleshy and convex labellum (vs. broadly obovate or rhombic-obovate when expanded) and its linear-obovate petals, erect, and directed upwards, parallel to the dorsal sepal (Dodson 1980, Vázquez & Dodson 1982).
The distribution known for this species has been from Venezuela to Bolivia, from 800 m (Dodson 1980) to 2700 m (Schweinfurth 1960) in elevation. Here it is reported at 350 m, expanding its geographical and elevational range.
Phalaenopsis Blume
Phalaenopsis stuartiana Rchb.f., Gard. Chron., n.s. 16(415): 748. 1881.
TYPE: [Philippines]. “Tropical Asiatic plant. Named in July last in honour of Mr. Stuart Low, of the old firm of Hugh Low & Co., is now in flower at Upper Clapton”, S. Low s.n. (holotype, W-R). Homotypic synonym: Phalaenopsis schilleriana var. stuartiana (Rchb.f.) Burb., Garden (London 1871-1927) 22: 119. 1882.
Heterotypic synonyms: Phalaenopsis schilleriana var. vestalis Rchb.f., Gard. Chron., n.s., 17: 330. 1882. Phalaenopsis schilleriana subvar. vestalis (Rchb.f.) A.H.Kent in H.J.Veitch, Man. Orchid. Pl. 7: 37. 1891. TYPE: The Philippines, hort. Low. s.n. (holotype, W). Phalaenopsis schilleriana var. alba Roebelen, Gard. Chron., ser. 3, 7: 459. 1890, TYPE: The Philippines, hort. Low. s.n. (holotype, W).
Description: Based on F. Pupulin 9082.
An epiphytic, fan-shaped, pseudobulbless, pendent herb to 22 cm tall excluding the inflorescence. Roots thick, flexuous, glabrous, distinctly flattened where adhering to the substrate, 3-7 mm in diameter, silvery white with orange-brown vegetative tips, occasionally forming plantlets on the upper surface. Stem short, the upper part completely enveloped by the sheathing bases of the leaf, the base sometimes exposed, about 2 cm wide, partially covered by the papyraceous, dry-fibrous, brownish white remnants of the leaf-sheaths. Leaves elliptic to oblong-elliptic, obtusely unequally bilobed, pendent, softly coriaceous to elastic, reddish green mottled with silvery gray, purple on the underside, 8.0-17.0 (40) × 3.3-5.2 cm, narrowed at the base into a short, broad conduplicate petiole less than 1 cm long, articulate with the conduplicate, short leaf-sheath. Inflorescence lateral, exerted from the axil of the lower leaves, a simple or paniculate, few- (2) to many- (over 100) flowered raceme, borne erect and becoming arched-pendent at flowering, distinctly longer than the leaves, 17-30 (80) cm long; peduncle terete, slender, purplish brown, to 25 cm long, completely covered by several tightly amplectent, triangular, obtuse, papyraceous, brown bracts to 3 × 3 mm; rachis slightly fractiflex. Floral bract papyraceous, pale brown, triangular, acute, ca. 5 × 3 mm. Pedicellate ovary terete, glabrous, gently curved, white, 3.0-3.6 cm long including the pedicel. Flowers spreading, faintly scented, the sepals and petals white, the labellar half of the lateral sepals pale greenish yellow spotted and blotched with brownish red, the lip white with base of the lobes yellow spotted and blotched with brownish red; the column white, the anther white flushed rose. Dorsal sepal elliptic to narrowly obovate, obtuse, 2.3-2.7 × 1.2-1.5 cm. Lateral sepals asimmetrically lanceolate, obtuse to subacute, 2.4-2.6 × 1.5-1.8 cm, the basal labellar margin reflexed to allow the insertion of the column foot between the sepals. Petals rhombic from a narrow cuneate base, obtuse-rounded to subtruncate, narrowly lanceolate-subfalcate, subacuminate, porrect, the apex facing down, 2.4-2.7 × 2.0-2.2 cm. Lip 3-lobed,1.8-2.0 cm long, 1.6-1.8 cm wide across the lateral lobes; lateral lobes oblong, rounded, erect, ca 1.0 × 0.5 cm; midlobe rounded, with a low longitudinal keel, ending into 2 narrowly triangular, digitiform-falcate, acuminate, retrorse extensions (cirrhi) 7 mm long, 1 mm wide at the base; the disc provided with a umbonate, erect, bilobed, high callus ca. 3 × 3 mm, the two halves elliptic, extending toward the rear into a short, pointed apicule ca. 1 mm long. Column semiterete subclavate, fleshy, 1 cm long, dilated in the middle around the stigma, extended at the base into a narrow, cuneate foot ca. 8 mm long; the rostellum a triangular, biparted teeth; the clinandrium triangular, shallow, entire, with a lower rim; the stigma ventral, broadly rounded, deep; the anther incumbent. Anther cap deeply cucullate, semi-ovoid, obtuse, 2-celled, the cells protected by subcircular, membranous flaps. Pollinarium composed of two pollinia, ovoid, hard, deeply cleft, bright orange, attached by small, lenticular caudicles to a triangular, attenuate, basally truncate, hyaline stipe apically adnate to the cordate-peltate, hyaline visicidium. Fruit not seen.
Eponymy: Named in honor of Mr. Stuart Henry Low (1826-1890), of the firm of Hugh Low & Co., at Upper Clapton, England, where the species flowered for the first time in Europe.
Distribution: Native from the island of Mindanao in the Philippines, the species has become naturalized in tropical America, where it has been recorded from the Atlantic coasts of Costa Rica and Panama (Central America), and Suriname (South America).
Habitat in Costa Rica: So far known exclusively from the wet tropical forests of the Caribbean plains, close to the border with Panama, where naturalized populations have been observed growing in secondary vegetation.
Phenology: Flowering in February and March.
Costa Rican material examined. Limón: Talamanca, Cahuita, west of Cahuita National Park, between Cahuita and Puerto Vargas, Hone Creek, vicinities of Cacao Trails Tayku, ca. 70 m, growing epiphytically (spontaneous) on the trunk of Cinnamomum sp. (Cinnamon) tree (Lauraceae), tropical rain forest, secondary vegetation with remnant trees of primary forest, flowered in cultivation and prepared 7 Mar. 2023, F. Pupulin 9082 & A. Mesén (JBL) (Fig. 18).

Native from the lowlands of the island of Mindanao in the Philippines, where it is endemic, Phalaenopsis stuartiana has been anecdotally known for a long time from the Caribbean coast of western Panama, where it escaped from cultivation, and it reproduces from seed in the wild. It was reported from Isla Colón in Bocas del Toro Archipelago (Bogarín et al. 2015) and, apparently, from the mangrove vegetation in Bocas del Toro lowlands (W. M. Whitten 3332, FLAS; A. Maduro & E. Olmos 251, MO 5892639). Only recently, however, photographs of wild specimens of P. stuartiana were photographed in situ (Hoskovec 2015) (Fig. 19), and the species was formally documented from a specimen collected from a natural population in Bocas del Toro Archipelago (Z. Sarracín, pers. comm. 2019). At least another wild collected plant of P. stuartiana is known from the Atlantic coast of South America, where B. B. Teunissen collected it in 1971 at Paramaribo, Suriname, in the framework of a survey carried out by the Suriname Forest Service (Teunissen LBB12998, U, with flowers in spirit). This is the first record of a naturalized population of P. stuartiana naturalized in Costa Rica. The plant was originally collected by Armando Mesen in hills close to Cahuita, where it belonged to a group of more than 20 plants living epiphytically on several close trees. Flowering in the wild has been recorded in February to May.

Phalaenopsis stuartiana Rchb.f. photographed in situ at Bocas del Toro, Panama, on 7 March 2014.
Pleurothallis R.Br.
Pleurothallis trigyna Pupulin, sp. nov.
TYPE: Costa Rica. San José: Pérez Zeledón, Páramo, Berlín, Fila Temblor, ca. 4 km north of Berlín, road towards the highest part of the Fila, 1807 m, rain premontane forest, epiphytic in secondary vegetation, 4 March 2011, flowered in cultivation and prepared 14 Sep. 2021, D. Bogarín 8427 et al. (holotype, JBL) (Fig. 20).

Diagnosis: Inter speciebus Pleurothallorum sectionis Macrophyllarum-Fasciculatarum, plantae robustae floribus flavis pallentibus labello bicolori, basi rosea pallente apice flavo fulgido, minutae guttulae transparentibus in basi labelli munitis, facile distinguenda.
Epiphytic, tall, caespitose herb, to about 20 cm tall. Roots slender, flexuous, ca. 1.5 mm in diameter. Ramicauls 11-18 cm long, enclosed by 2 tubular, obtuse, tightly adpressed, papyraceous, brown sheaths, the first one basal, ca. 2 cm long, the second inserted in the lower third of the ramicaul, 4-5 cm long. Leaves coriaceous, pale green, ovate or lanceolate, abruptly acute, the apex minutely excised, 6-8 × 5-6 cm, deeply cordate at the conduplicate base, the basal margins not overlapping, the midvein slightly decurring on the stem. Inflorescence with single-flowered coflorescences, exerted from a narrow rectangular, obliquely acute, brown, papery, prostrate spathaceous bract 1.5 cm long; pseudopeduncle filiform, ca. 1 cm long. Floral bract triangular-ovate, subacute, glumaceous, to 2 mm long. Pedicel terete, to 2.5 mm long. Ovary terete-subclavate, rounded, stout, to 2.5 mm long. Flowers bilabiate, not completely spreading, with the sepals pale yellow, the petals pale yellow faintly suffused with rose at the base, the lip yellow from a pale rose base, the column pink from a greenish white base, the anther white, flushed pink at the base. Dorsal sepal oblong, obtuse, abruptly acutiuscule at the apex, with a rounded apicule, 7 × 4 mm, 5-veined. Lateral sepals fused into an ovate, obtuse, rounded-apiculate, 7-veined synsepal, 7.0 × 5.5 mm. Petals ligulate-down curved, acute with the apex minutely rounded, 4.5 × 1 mm, 1-veined, the base thickened into a low, semircircular callus. Lip three-lobed, subrectangular-subpandurate, concave, 3.5 × 2.7 mm, the base subtruncate with a central, semicircular depression, the lateral lobes rectangular-elliptic, papillose, the papillae decurring to the base of the midlobe, the median lobe transversely hemielliptic-ovate, truncate, ca. 2.0 × 2.5 mm; disc with a rounded glenion ca. 0.5 × 0.5 mm, flanked by subacute, protruding calli. Column short, thick, conical, dorsiventrally flattened, 1.5 mm long, the clinandrium 3-toothed, the anther dorsal, the stigma apical, bilobed, with two parastigmatic, triangular, acuminate lobes ending in spherical viscidia. Anther cap lanceolate, cucullate, minutely papillose, 2-celled. Pollinarium composed of two fusiform pollinia, apically attenuate-recurved, and a spherical viscidium. Fruit not seen.
Etymology: From the Greek words τρία (tría), three, and γυνή (gunḗ), female, in allusion to the three rostella and viscidia, which are produced from the stigmatic lobes of the gynostemium.
Distribution: Only known from Costa Rica.
Ecology: The species is known to inhabit the rain and wet premontane and lower montane forests of the Pacific drainage in the Cordillera de Talamanca, at elevations around 1800 m.
Phenology: Flowering from June to October.
Distinguishing features: Pleurothallis trigyna can be recognized by the comparatively tall plants for its group, reaching about 20 cm, the very coriaceous-thickened leaves, the solitary flowers not completely spreading, pale yellow, with a pandurate, bicolored lip (basally pale rose, apically bright yellow), the small, rounded glenion flanked by subacute, protruding calli, and the production of a large amount of transparent droplets at the base of the lip. All the flowers of the type specimen, even in different flowering times, are characterized by a gynostemium producing three rostella, each one topped by a spherical viscidium.
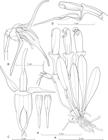
During the preparatory work aimed at a systematic revision of the small-flowered Pleurothallis of sect. Macrophyllae-Fasciculate (Pupulin et al. in prep.), one of the most curious novelties to appear was the species proposed here as P. trigyna. It belongs to a group of still unpublished species that mostly inhabit premontane and montane forests close to 2000 m elevation. The group is characterized by rather large plants with fleshy leaves and small, often ringent flowers, provided with a rectangular to pandurate lip. Close to its base and around the glenion, the ventral surface of the lip produces a copious number of droplets made of a clear, transparent substance that we have not determined yet as to its chemical properties. Other examples of species in this group are the specimens M. Díaz 237, A.P. Karremans 6077, D. Bogarín 9091, and D. Bogarín 8739 (all vouches in JBL), which in all probability belongs to another four different taxa (Fig. 21). Among them, P. trigyna may be recognized by the pandurate lip, unique in species of Macrophyllae-Fasciculatae in Costa Rica, which is pale rose at the base and bright yellow at the apex. The unique characteristic of the specimen used to prepare the holotype of the new species is that the column presents three rostella, each one ending into a spherical viscidium. Only one of the viscidia is completely functional, as it is associated with a fertile anther, from which the apexes of the two pollinia elongate until they become connected to the viscidium, making the pollinarium available for removal by a pollinator (Fig. 22). As we do have only this single specimen available for study, we refrain from consider this feature as characteristic of the species per se, but nonetheless all the flowers of the specimen Bogarín 8427 present a trigynous column, indicating that this character is genetically controlled.


Column of Pleurothallis trigyna Pupulin showing the three rostella with viscidia, only one of which associate to a fertile anther.
Specklinia Lindl.
Specklinia echinata (L.O.Williams) Soto Arenas & Solano, Icon. Orchid. (Mexico) 5-6: t. 670. 2002 (2003). Bas. Pleurothallis fuegi var. echinata L.O.Williams, Ann. Missouri Bot. Gard. 33(1): 120. 1946.
TYPE: Panama. Chiriquí: Volcán de Chiriquí, alt. about 2720 m, 15 Jul. 1938, M.E. Davidson 981 (holotype, AMES!).
Description: Based on Karremans 5600 and Karremans 8822 & I. Chinchilla.
Plant small, epiphytic, caespitose, up to 5 cm tall including the inflorescence. Roots slender, 1 mm wide. Ramicauls erect, minute, up to 5-6 mm long, enclosed by 2 thin tubular sheaths. Leaf erect, coriaceous, obovate to broadly elliptic, gradually narrowed into a petiolate base, obtuse, apiculate, apically bilobed, 1.2-1.5 × 0.5-0.6 cm. Inflorescence with erect, thin, loose, sub-secund, successively muti-flowered flowered coflorescences, up to 4.5 cm long including the pseudopeduncle up to 3.5 cm long; born from the apex of the ramicaul. Floral bracts imbricate, involving the ovary and rachis, membranaceous, apiculate, 1 mm, pedicel cylindrical, bent, ovary 0.8 mm long, tricarinate, covered in transparent filiform glands. Flowers dimorphic, transparent white with purplish veins and lip or yellow with purplish veins. Sepals membranaceous, carinate, glabrous, the dorsal sepal elliptic-oblong, 7.5-9.2 × 2.3-2.5 mm, 3-veined, acute, contracted into a thickened narrow tail. Lateral sepals connate to near the middle into a bifid synsepal, each lateral sepal ovate, 7.5-9.5 × 1.8-2.0 mm, 2-veined, acute, narrowing into a thickened tail. Petals oblanceolate, obtuse, rounded at the apex, 2.9-3.0 × 0.9-1.1 mm, 1-veined. Lip oblong, subtrilobed, rounded-truncate apically, 2.7-3.8 × 1.6-1.8 mm, with a pair of median elevated lobes, delicately hinged to the column foot. Column with a pair of subquadrate apical lobes, 2 mm long, the foot as long as the body, stigma ventral. Anther cap incumbent, cucullate, white. Pollinarium composed of two pollinia, naked flattened. Fruit not seen.
Etymology: From the Latin echinatus, “spiny like a hedgehog”, alluding to the ovary.
Distribution: Known to occur from Mexico to Panama.
Habitat in Costa Rica: Epiphytic on the Pacific watershed of the Cordillera de Talamanca growing in primary oak forest from 2300 to 2500 m.
Phenology: Flowering between June and August.
Costa Rican material studied: San José: Dota, Distrito de Copey, Parque Nacional Los Quetzales, 3 km sobre el camino a Providencia desde Ojo de Agua, 2500 m, bosque nuboso montano, 4 ago. 2012, A.P. Karremans 5600 (JBL-D6073 spirit!; Fig. 23). Dota, Distrito de Copey, Parque Nacional Los Quetzales, 3.5 km sobre el camino a Providencia desde Ojo de Agua, 2300 m, bosque nuboso montano, 25 jun. 2021, A.P. Karremans 8822 & I. Chinchilla (JBL-A0780 spirit!; Fig. 24).


Luer (2006) treated Specklinia echinata as a synonym of S. fuegi. Soto Arenas & Solano (2003) separated the two names based on the larger plants and flowers, reddish, less arcuate, not so neatly 3-lobed lip and shorter claw. Based on the original illustration and description by Reichenbach, it seems that S. fuegi has a glabrous ovary and shorter, thicker tails, whereas S. echinata has an echinate ovary and long, narrow tails. We follow Karremans et al. (2016) in recognizing the two taxa as distinct. Both S. echinata and S. fuegi have been reported to occur in Mexico, Guatemala, Honduras, El Salvador, Nicaragua and Panama, and although Karremans & Vieira-Uribe (2020) included a photo of the latter from Costa Rica, neither had been formally recorded for the country’s flora with a voucher. Here S. echinata is described and illustrated based on materials collected on the Pacific slope of the Cordillera de Talamanca. In Costa Rica, plants with smaller yellow flowers grow on the same branch, with plants bearing larger whitish with purple flowers. No specimens agreeing with S. fuegi (sensu Soto Arenas & Solano 2003) have been recorded for the country.
This species belongs to Specklinia subgen. Sylphia (Luer) Karremans, a little group of five species which in Costa Rica is represented by S. absurdaBogarín, Karremans & Rincón, S. echinata and S. turrialbae (Luer) Luer (Bogarín et al. 2013, Karremans et al. 2016).
Stelis Sw.
Stelis veraguasensisLuer, Selbyana 22(2): 126. 2001.
TYPE: Panama. Veraguas: alt. ca. 500 m, 5 Sep. 1976, C. Luer & R. Dressler 1235 (holotype, SEL-001096).
Description: Based on D. Bogarín 10490
Plant epiphytic, caespitose, erect, up to 32 cm tall. Roots slender, flexuous, up to 3.5 cm long, 2 mm in diameter. Ramicauls erect, stout, 6-15 cm long, enclosed by 2-3 loose, tubular, obtuse sheaths, 3-5 cm long. Leaf elliptic, obtuse, coriaceous, green, 13-18 × 2-5 cm, including the cuneate base narrowing into a petiole 1-2 mm long. Inflorescence with erect, compact, distichous, multi-flowered coflorescences, with several flowers open simultaneously, up to 22.5 cm long including the pseudopeduncle 7.8 cm long, from an annulus below the apex of the ramicaul, rachis up to 13.8 mm long. Floral bracts greenish to pale beige with age, infundibular, obtuse, 0.3-0.7 mm long. Ovary ca. 3 mm long, pedicel 3-4 mm long. Flowers bilabiate, the sepals cream, slightly tinged with fuchsia, the petals fuchsia with a light yellowish-green hue, vinous at the base, the lip vinous, edged in yellowish-green, the yellowish-green, lustrous glenion, the column vinous. Dorsal sepal elliptic, obtuse, convex, microscopically papillose, margins microscopically ciliate, connate to the lateral sepals for about 3 mm, 12 × 12 mm, 7-nerved. Lateral sepals asymmetrically ovate, obtuse, connate for 10 mm into a broadly ovate broadly ovate, concave synsepal, 11 × 13 mm, 9-nerved. Petals broadly overlapping above the column, ovate, cordate, acute, the base narrowly concave, 2.5 × 2.8 mm, 1-nerved. Lip ovate, truncate, obtuse, thickened along the margin, the base transversely thickened with a large glenion 1.1 × 0.6 mm, lanceolate, obtuse in contour, hinged to the base of the column, 1.6 × 1.8 mm, 3-nerved. Column porrect, stout, non-foot, 1.3 × 1.7 mm, with the anther apical, the stigma apical, bilobed. Anther cap, ovate, abaxially concave, 0.20 × 0.15 mm. Pollinarium composed of two pollinia, ovoid, ca. 0.10 × 0.07 mm, joined at the base by a rounded viscidium. Fruit not seen.
Etymology: Named for Veraguas, the Panamanian province where the type specimen was collected.
Distribution: The species is known from Costa Rica and Panama.
Habitat in Costa Rica: Plants were found growing as epiphytes on the Pacific basin lower montane rain forest, west of the Talamanca Mountain Range, in Las Tablas Protective Zone at 1778 m.
Phenology: Flowered in November under cultivation.
Costa rican material studied: Puntarenas: Coto Brus, Sabalito, Zona Protectora Las Tablas, 13 km al noreste de Lucha, sitio Coto Brus, Finca de Miguel Sandí, 1778 m., floreció en cultivo en el Jardín Botánico Lankester, preparado 19 nov. 2013, D. Bogarín 10490 &D. Jiménez (JBL-D6150 spirit!, Fig. 25-26).


Stelis veraguasensis is a large, robust plant with a long, multiflowered raceme of large, bilabiate, shell-shaped flowers that open simultaneously. According to Luer (2001), it is similar to S. lankesteri Ames, a species from Costa Rica and Panama. However, S. veraguasensis is distinguished by having a larger habit (ca. 32 cm vs. 20-22 cm) with generally broader leaves (4.0-5.5 cm vs. 2.0-2.5 cm), larger sepals (11-12 × 12-13 mm vs. 8.0-9.0 × 7.0-7.5 mm), larger petals (2.5 × 2.8 mm vs. 2.0 × 2.0 mm), ovate, cordate (vs. rhombic) and larger lip (1.6 × 1.8 mm vs. 1.0 × 2.0 mm), with a thick v-shaped apical margin, and a short, rounded apiculum.
Telipogon Kunth
Telipogon memoria-rodulfii Pupulin & Bogarín, sp. nov.
TYPE: Costa Rica. Cartago: Casamata del Guarco, 1930 m, epiphytic on short trees in pasture, mostly Quercus spp. (Fagaceae), montane cloud forest, 26 March 2002, F. Pupulin 3557 et al. (holotype, USJ). (Fig. 27, 28A).


Diagnosis: A Telipogon fortunae Dressler floribus duplo maioribus rosaceis brunneo striatis recedit.
Epiphytic, small, monopodial herb to 5 cm long, including the inflorescence. Roots flexuous, numerous, short, to 1.5 mm in diameter, with green apices. Stem abbreviated, completely concealed by the base of the amplectant leaf-sheaths. Leaves 2, elliptic, acute, gently ribbed on the underside, 4.8-6.6 × 1 mm, sometimes deciduous at flowering. Inflorescence lateral, from the base of the stem, a successively 2-3-flowered raceme to 5 cm long, sometimes with a branch about 2.5 cm long in the upper half; the peduncle born terete, slender, becoming dorsiventrally flattened-ancipitous toward the apex, the flattened internodes provided with a triangular, obtuse, weakly keeled, pale green, glumaceous bract at the apex, decurrent on the peduncle; the rachis subtrigonous, straight. Floral bract triangular, acute, rigid, strongly conduplicate-keeled, ca. 1.5 × 1.0 mm. Pedicellate ovary terete-subclavate, rounded in section, 2.5 mm long including the pedicel. Flowers non-resupinate, pale rose, the sepals darker, the petals and lip longitudinally striped with pale yellowish brown, the column purple, the anther cap red. Dorsal sepal triangular-lanceolate, acute, reflexed, the margins shortly inflexed, 3.5 × 1.4 mm, 3-veined. Lateral sepals adpressed, triangular-lanceolate, acute, lightly falcate at apex, 3.3 × 1.0 mm, 1-veined, the margins lightly involute to become adpressed to the apex of the lip. Petals lanceolate, acute, 8 × 3 mm, 3-veined. Lip lanceolate, acute, slightly reflexed, 6.2 × 2.4 mm. Column semiterete, 2.5 mm long, the stigma recessed under a long, narrowly triangular stigmatic lobe, the anther dorsal (facing down in natural position), the rostellum long-attenuated, dilated at apex, far surpassing the apex of the stigmatic lobe. Anther cap cucullate, cordate, acute, with two large flaps protecting the pollen. Pollinarium composed of four pollinia, discoidcomplanate, in two pairs of different size, on a triangular, conduplicate, hyaline stipe and a hook-shaped, brown viscidium. Fruit not seen.
Eponymy: From the Latin memoria, remembrance, and Rodulfus, Rudolf, in the memory of our dear friend, untimely passed away, Rudolf Jenny (1953-2021).
Distribution: Only known from Costa Rica.
Habitat in Costa Rica: The species is known from the Caribbean watershed of the northern portion of the Talamanca Cordillera in Costa Rica, where it inhabits the premontane rain and wet forest around the Tapantí National Park.
Phenology: Flowering in March, May, and October.
Costa Rican material studied: Cartago: Cartago, San Francisco, Muñeco, 4.5 km south of Muñeco, road to Alto Belén, 1968 m, premontane rain forest, epiphtic in secondary vegetation and trees in open areas, 27 May 2009, D. Bogarín 6582 et al. (USJ) (Fig. 28B). Paraíso, Orosi, Tapantí, Parque Nacional Tapantí, ca. 9 km after the gate of the view point, road to Tapantí Hydroelectric Project, ca. 100 m after the waterfall, 1694 m, premontane rain forest, epiphytic on twigs of Saurauia montana (Actinidiaceae), 26 Aug. 2021, D. Bogarín 13425 & G. Villalobos (JBL) (Fig. 28C).
Telipogon memoria-rodulfii is another species belonging to Stellilabium sect. Rhamphostele sensuDressler (1999). It is most similar to T. fortunae (Dressler) N.H.Williams & Dressler, but the latter has much smaller flowers (all the tepals approximately half in size), and the flowers are yellow or purple (Dressler 1999).
Trichopilia Lindl.
Trichopilia olmosiiDressler, Selbyana 22(1): 11-12. 2001.
TYPE: Panama. Bocas del Toro: region of Culebra, 1000-1200 m, collected by Erick Olmos; flowered in cultivation 4 Aug. 2000, R. L. Dressler 6288 (holotype, barcode MO-1005420!).
Description: Based on F. Pupulin 9118.
An epiphytic, caespitose, pseudobulbous herb to 17 cm tall. Roots slender, flexuous, glabrous, ca. 1.5 mm in diameter. Rhizome with short internodes, covered by brown, nervous, papyraceous sheaths. Pseudobulbs ovate-oblong, markedly compressed sublenticular, ancipitous, 3.3-4.8 × 1.5-2.1 cm, monophyllous, covered at the base by 2-3 triangular, acute, imbricating sheaths to 3.8 cm long, born glumaceous and becoming dry-papyraceous with age, eventually becoming lacerate and dissolving. Leaves oblong-elliptic, grass-green, acute to subobtuse, weakly keeled beneath, matte on both surfaces, 10-12 × 3-4 cm, the margins slightly undulated and sometimes recurved, narrowed at the base in a short, folded petiole to 2 cm long, ca 0.5 cm wide. Inflorescence a few-flowered (2-5), pendent raceme 6-10 cm long, covered near the base by an amplectent, somewhat loose, glumaceous, tubular bract to 1.7 × 0.5 cm, brownish spotted dark brown. Floral bracts glumaneous, greenish brown, narrowly ovate, acute, carinate, 1.4-1.9 × 0.9-1.2 cm. Pedicellate ovary terete-subclavate, with deep intercarpellar grooves, 2.4- 2.9 cm long including the pedicel. Flowers spreading, with greenish yellow to pale whitish-green spells, the lip white with bright yellow blotches on the callus and the osmophores. Dorsal sepal narrowly elliptic, acute, weakly carinate, 2.3-2.7 × 0.5-0.6 cm. Lateral sepals narrowly elliptic-subfalcate, short acuminate, weakly carinate, 2.5-2.7 × 0.4-0.5 cm, shortly connate at the base for about 5 mm. Petals elliptic-lanceolate, acute-subapiculate, carinate in the distal third, recurved to gently reflexed in natural position, 2.3-2.8 × 0.5-0.6 cm. Lip 3-lobed, broadly ovate from a cuneate base, 1.8-2.1 × 1.7-2.0 cm, basally connate to the column for ca. 5 mm, the disc provided with a distinct keel ca. 8 mm long, reaching the middle of the blade; lateral lobes rounded, erect, encircling the column; midlobe excise, divided in 2 distinct, rounded subquadrate lobes, with slightly waved margins. Column terete, abruptly dilated around the stigma, ca. 1.3 × 0.6 cm; the clinandrium cucullate, lacerate; the stigma ventral, broadly elliptic, large, with a distinct rim; the anther incumbent. Anther cap cucullate, ovate-subquadrate, obtuse, 2-celled. Pollinium composed of two pollinia, ovoid, slightly complanate, hard, deeply cleft, on a narrowly triangular, basally marginate, hyaline stipe and an elliptic, orange viscidium. Fruit not seen.
Costa Rican material examined. San José: Vázquez de Coronado-Moravia, Jesús-San Jerónimo, from Alto La Palma to Bajo La Hondura, ca. 10°02’N 83°59’W, 1500 m, premontane rain forest, flowered in cultivation at Jardín Botánico Lankester and prepared 21 Sep. 2023, F. Pupulin 9118 & G. Villalobos (JBL, Fig. 29).

Dressler (2001) noted that Trichopilia olmosii is morphologically similar to both T. maculata Rchb.f. from Panama, and the Costa Rican Trichopilia turialbae Rchb.f. Trichopilia maculata is easy to distinguish for its oblong-subquadrate pseudobulbs that are closely appressed to the substrate (Dressler 2001), as well as the single-flowered inflorescence, and even though this name has been applied to plants from Costa Rica, El Salvador, and Guatemala, we have never seen a true specimen of T. maculata in Costa Rica. We suspect that the species is indeed endemic to central Panama. Trichopilia turrialbae, and the similar T. endresiana Dressler & Pupulin, have much larger flowers, with lateral sepals connate to about the half of their length, and the lip of both species is distinctly convex in lateral view, unlike the almost straight lip of T. olmosii.
Additions and corrections regarding the orchid flora of Costa Rica
Omissions and taxa described after the publication of the most recent Costa Rican orchid catalogue (Pupulin et al. 2023) are discussed here. The orchid flora of Costa Rica now includes 1695 species and nine forms.
Funkiella valerioi (Ames & C.Schweinf.) Salazar & Soto Arenas, Acta Bot. Mex. 97: 52. 2011.
Bas.: Spiranthes valerioi Ames & C.Schweinf., Schedul. Orchid. 10: 8. 1930.
Spiranthes parasitica var. valerioi (Ames & C.Schweinf.) L.O.Williams, Ceiba 1: 186. 1950.
Schiedeella valerioi (Ames & C.Schweinf.) Szlach. & Sheviak, Rhodora 92: 16. 1990.
Voucher: P.C. Standley & J. Valerio 43952 (AMES).
Note: Inadvertently, Funkiella valerioi (Ames & C.Schweinf.) Salazar & Soto Arenas and Schiedeella valerioi (Ames & C.Schweinf.) Szlach. & Sheviak were listed as accepted names. However, they refer to the same species. The accepted name here is F. valerioi.
Karma pusilla (Kunth) Karremans, Harvard Pap. Bot. 28: 67. 2023.
Dendrobium peruvianum F.Dietr. in Neu. Nachtr. Vollst. Lex. Gärtn. 3: 354. 1834., nom. superfl.
Dendrobium pusillum Kunth in F.W.H.von Humboldt, A.J.A.Bonpland & C.S.Kunth, Nov. Gen. Sp. 1: 357. 1816.
Humboltia pusilla (Kunth) Kuntze in Revis. Gen. Pl. 2: 668. 1891.
Pleurothallis pusilla (Kunth) Lindl. in Edwards’s Bot. Reg. 28(Misc.): 82. 1842.
Specklinia pusilla (Kunth) Lindl. in Edwards’s Bot. Reg. 21: t. 1797. 1835.
Trichosalpinx pusilla (Kunth) Luer in Phytologia 54: 397. 1983.
Tubella pusilla (Kunth) Archila in Revista Guatemalensis 3(1): 62. 2000 publ. 2009.
Voucher: M. Fernández et al. 911 (JBL-spirit).
Note: This species was previously recorded for Costa Rica but omitted in the catalogue (Pupulin et al. 2023). Here we cite a Costa Rican voucher.
Karma todziae (Luer) Karremans, Harvard Pap. Bot. 28: 68. 2023.
Trichosalpinx todziaeLuer in Lindleyana 11: 111. 1996. Tubella todziae (Luer) Archila in Revista Guatemalensis 3(1): 66. 2000 publ. 2009.
Voucher: C. Todzia 344 (CR).
Note: This species was described from Costa Rica but omitted in the catalogue (Pupulin et al. 2023).
Malaxis dresslerianaChinchilla, Karremans & M.A.Blanco, Orchids (West Palm Beach) 92: 609. 2023.
Voucher: I. Chinchilla et al. 3209 (JBL-spirit).
Note: This species was described after the publication of the catalogue (Pupulin et al. 2023).
Maxillariella ponerantha (Rchb.f.) M.A.Blanco & Carnevali, Lankesteriana 7: 529. 2007.
Maxillaria ponerantha Rchb.f., Bonplandia (Hannover) 2: 17. 1854.
Voucher: P.H. Allen 5328 (SEL).
Note: This species was previously documented in Costa Rica but was unintentionally excluded from the catalogue (Pupulin et al. 2023). Although the populations in Costa Rica differ from the original type specimen based on Venezuelan material (Wagener s.n., W), we treat this species under M. ponerantha until this taxonomic concept is thoroughly reviewed.
Sarcoglottis woodsonii (L.O. Williams) Garay, Bot. Mus. Leafl. 28(4): 355. 1980[1982].
Voucher: R. Acuña et al. 3282 (USJ, JBL-spirit).
Note: This species was first documented in Costa Rica by Acuña-Castillo et al. (2024).
Telipogon fortunae (Dressler) N.H.Williams & Dressler, Lankesteriana 5: 169. 2005.
Voucher: I. Chinchilla et al. 3209 (JBL-spirit).
Note: This species was previously recorded for Costa Rica but omitted in the catalogue (Pupulin et al. 2023). Here we cite a Costa Rican voucher.
Telipogon lateritius Pupulin, Lankesteriana 24(1): 71-73. 2024.
Voucher: F. Pupulin et al. 4186 (USJ).
Note: This species was described after the publication of the catalogue (Pupulin et al. 2023).
Telipogon muntziiBogarín, O.Pérez & Pupulin, Lankesteriana 24(1): 73-76. 2024.
Voucher: D. Bogarín et al. 14185 (JBL-spirit).
Note: This species was described after the publication of the catalogue (Pupulin et al. 2023). The epithet honors Dr. Robert Muntz. In the eponymy, the surname was originally stated as Robert Müntz (Bogarín et al. 2024), but it should be spelled as Muntz without the umlaut.
Trichosalpinx orbicularis (Lindl.) Luer, Phytologia 54: 396. 1983.
Humboltia orbicularis (Lindl.) Kuntze in Revis. Gen. Pl. 2: 668. 1891.
Pleurothallis orbicularis (Lindl.) Lindl. in Edwards’s Bot. Reg. 28(Misc.): 79. 1842.
Specklinia orbicularis Lindl. in Edwards’s Bot. Reg. 25(Misc.): 31. 1839.
Voucher: M. Fernández et al. 65 (JBL-spirit)
Note: This species was previously recorded for Costa Rica but omitted in the catalogue (Pupulin et al. 2023). Here we cite a Costa Rican voucher.
Acknowledgements
We are thankful to the Costa Rican Ministry of Environment and Energy (MINAE) and the National System of Conservation Areas (SINAC) for issuing the Scientific Passports that allowed the collection of wild specimens treated in this study. We acknowledge Steve Kirby, Lyle Rice, and the González-Sotela family for cofunding the Bosque de Paz Orchid Conservation Project, through which one taxon included in this treatment was found. We are grateful to Sara Díaz Poltronieri, illustrator at JBL, for her collaboration on the Lepanthes acarina illustration. Jindřiška Vančurová kindly provided permission for the reproduction of their photos. We also thank the personnel at CR, JBL, and USJ for granting full access to their collections. Special thanks to Leonardo Álvarez, Coordinador de Conservación Botánica of Osa Conservation, for sharing material and information. Finally, we appreciate the valuable recommendations from three anonymous reviewers.
Funding: The Vice-Presidency of research of the University of Costa Rica provided support under the projects “Inventario y taxonomía de la flora epífita de la región Mesoamericana” (814-A7-015), “Flora Costaricensis: Taxonomía y Filogenia de la subtribu Pleurothallidinae (Orchidaceae) en Costa Rica” (814-BO-052), “Filogenia molecular de las especies de Orchidaceae endémicas de Costa Rica” (814B1-239), “La flora de orquídeas del Monumento Nacional Guayabo, Turrialba, Costa Rica” (814-B5-115), “Hacia una moderna flora de orquídeas de Panamá” (814-B2-161), and “Taxonomía de Telipogon s.l. and Ornithocephalus in Costa Rica: addenda to Oncidiinae” (C0051).
Author contributions: NBO conceptualized the study, compiled information, wrote the original draft, and contributed to the species descriptions, reviewing, editing, LCDP design, and figure standardization. GS, LO, KGA, GRA, and IFC participated in species description, LCDPs and illustrations design, writing, reviewing, and editing. MDM contributed to conceptualization and data compilation, writing, reviewing, and editing. FP, DB, and APK participated in conceptualization, species description, writing, reviewing, editing, LCDP and illustration design, and manuscript supervision.
Conflict of interest: The authors declare no conflicts of interest.
Literature cited
- Acuña-Castillo, R., Blanco, M.A., Artavia, M., Jiménez, J.E. & Bogarín, D. (2024). Sarcoglottis woodsonii (Orchidaceae: Spiranthinae)-rediscovered in Costa Rica after 80 years, with a preliminary survey on aquatic and other wetland orchids. Harvard Papers in Botany, 29(1), 15-33.
- Atwood, J. T. & Mora De Retana, D. E. (1999). Family #39, Orchidaceae: Tribe Maxillarieae: Subtribes Maxillariinae and Oncidiinae. In: W. Burger (Ed.), Flora Costaricensis. Fieldiana, Botany, n.s. 40.
- Barros, F. & Barberena, F. F. (2010). Nomenclatural notes and new combinations on Anathallis and Specklinia (Orchidaceae). Rodriguésia, 61(1), 127-131.
- Bernal, R., Gradstein, S. R. & Celis, M. (eds.). (2019). Catálogo de plantas y líquenes de Colombia Bogotá: Instituto de Ciencias Naturales, Universidad Nacional de Colombia.
- Blanco, M. A. (2014). Four new species of Lockhartia (Orchidaceae, Oncidiinae). Phytotaxa 162(3), 134-146.
- Blanco, M. A., Jiménez, J. E. & Juárez, P. (2016). Mormodes salazarii (Orchidaceae, Catasetinae), a new species with greenish-white flowers from Costa Rica. Phytotaxa, 245(2), 161-168.
- Bogarín, D. (2015). A new Campylocentrum (Orchidaceae, Angraecinae) from Parque Internacional La Amistad, Costa Rica. Novon, A Journal for Botanical Nomenclature, 24(2).
- Bogarín, D. (2020). A new species of Eurystyles (Orchidaceae: Spiranthinae) from Costa Rica. Blumea, 65(1), 65-68.
-
Bogarín, D. & Karremans, A. P. (2015). Prosthechea tinukiana sp. nov. (Orchidaceae: Laeliinae): an update of the Prosthechea prismatocarpa complex. Turkish Journal of Botany, 39(3), 499-505. https://doi.org/10.3906/bot-1408-38
» https://doi.org/https://doi.org/10.3906/bot-1408-38 - Bogarín, D. & Karremans, A. P. (2016). A new Brachionidium (Orchidaceae: Pleurothallidinae) from the first botanical expedition to the Volcán Cacho Negro, Costa Rica. Systematic Botany, 41(4), 919-923.
- Bogarín, D. & Kisel, Y. (2014). A new Lepanthes from Costa Rica. Orchid Review, 122(1305), 28-31.
- Bogarín, D. & Pupulin, F. (2010). Two new species of Mormolyca from Costa Rica and Panama. Orchid Digest, 74(1), 43-47.
- Bogarín, D. & Pupulin, F. (2021). The Orchid Flora of Barra Honda National Park, Nicoya, Guanacaste, Costa Rica. Harvard Papers in Botany, 26(1), 7-99.
- Bogarín, D., Chinchilla, I. F. & Cedeño-Fonseca, M. (2020). Two new species of Lepanthes (Orchidaceae: Pleurothallidinae) from Costa Rica and their phylogenetic affinity. Plant Systematics and Evolution, 306, 20.
- Bogarín, D., Kaes, E., & Díaz-Morales, M. (2019). Lepanthes elusiva - a new species of Lepanthes (Orchidaceae: Pleurothallidinae) from Tapantí Area in Cartago, Costa Rica, and a glance to other species of the genus in a small area around the Río Grande de Orosi in Costa Rica. Die Orchidee, 5(03), 19-28.
- Bogarín, D., Karremans, A. P. & Muñoz, M. (2015). Brachionidium kirbyi, a new species honoring the founder of the “Bosque de Paz” Orchid Project in Costa Rica. Die Orchidee, 66(5), 404-409.
- Bogarín, D., Karremans, A. P & Pupulin, F. (2008). New species and records of Orchidaceae from Costa Rica. Lankesteriana, 8(2), 53-74.
- Bogarín, D., Karremans, A. P., Rincón, R. & Gravendeel B. (2013). A new Specklinia (Orchidaceae: Pleurothallidinae) from Costa Rica and Panama. Phytotaxa, 115(2), 31-41.
- Bogarín, D., Oses, L. & Smith, C. M. (2017). Masdevallia luerorum (Orchidaceae: Pleurothallidinae), a new species from Costa Rica. Lankesteriana, 17(2), 235-244.
- Bogarín, D., Pérez-Escobar, O. A., Groenenberg, D., Holland, S. D., Karremans, A. P., Lemmon, E. M., Lemmon, A. R., Pupulin, F., Smets, E. & Gravendeel, B. (2018a). Anchored hybrid enrichment generated nuclear, plastid and mitochondrial markers resolve the Lepanthes horrida (Orchidaceae: Pleurothallidinae) species complex. Molecular Phylogenetics and Evolution, 129, 27-47.
-
Bogarín, D., Pupulin, F. & Pérez-Escobar, O. A. (2024). Two new species of Telipogon (Oncidiinae) from Costa Rica. Lankesteriana, 24(1), 69-77. http://dx.doi.org/10.15517/lank.v24i1.59556
» https://doi.org/http://dx.doi.org/10.15517/lank.v24i1.59556 - Bogarín, D., Pupulin, F., Smets, E. & Gravendeel, B. (2016). Evolutionary diversification and historical biogeography of the Orchidaceae in Central America with emphasis on Costa Rica and Panama. Lankesteriana, 16(2), 189-200.
- Bogarín, D., Serracín, Z. & Samudio, Z. (2014). Illustrations and studies in Neotropical Orchidaceae. The Specklinia condylata group (Orchidaceae) in Costa Rica and Panama. Lankesteriana, 13(3), 185-206.
- Bogarín, D., Serracín, Z. & Samudio, Z. (2018). A new Maxillariella (Orchidaceae: Maxillariinae) from Costa Rica and Panama. Brittonia, 70, 394-398.
- Bogarín, D., Serracín, Z., Samudio, Z., Rincón, R., & Pupulin, F. (2015). An updated checklist of the Orchidaceae of Panamá. Lankesteriana, 14(1), 135-364.
- Chinchilla, I. F., Aguilar, R. & Bogarín, D. (2020). A new Lepanthes (Orchidaceae: Pleurothallidinae) from Península de Osa, Puntarenas, Costa Rica. Harvard Papers in Botany, 25(2), 215-219.
-
Chinchilla, I. F., Karremans, A. P. & Blanco, M. A. (2022). A new species and a new record of Malaxis (Malaxidinae) from Costa Rica. Lankesteriana, 22(1), 37-51. 10.15517/lank.v22i1.50848
» https://doi.org/10.15517/lank.v22i1.50848 -
Chinchilla, I. F., Karremans, A. P. & Blanco, M. A. (2023). Malaxis dressleriana: A new orchid species honoring a botanist for all seasons. Orchids, 92(8), 608-613. 10.15517/lank.v22i1.50848
» https://doi.org/10.15517/lank.v22i1.50848 - Christenson, E., Harding, P. A., McIllmurray, M. & Blanco, M. A. (eds.) (2013). Maxillaria An unfinished monograph. Lebanon, Oregon: Privately published by Patricia A. Harding for Robert Christenson.
- Díaz-Morales, M. & Karremans, A. P. (2016). Epidendra Nova Talamancana. Phytotaxa, 272(4), 248-256.
- Díaz-Morales, M. & Pupulin, F. (2019). The New Refugium Botanicum. Phragmipedium × talamancanum. Orchids
- Dodson, C. H. & Dodson, P. M. (1980). Orchids of Ecuador. Icones Plantarum Tropicarum, 2, i-ii, t. 101-200.
- Dormont, L., Joffard, N. & Schatz, B. (2019). Intraspecific variation in floral color and odor in orchids. International Journal of Plant Sciences, 180(9), 1036-1058.
- Dressler, R. L. (1999). A reconsideration of Stellilabium and Dipterostele. Harvard Papers in Botany, 4(2), 469-473.
- Dressler, R. L. (2001). Mesoamerican orchid novelties 5, Oncidiinae. Selbyana, 22, 9-13.
- Dressler, R. L. (2003). Orchidaceae. En: B.E. Hammel, M.H. Grayum, C. Herrera & N. Zamora (eds.), Manual de Plantas de Costa Rica. Vol. III. Monographs in Systematic Botany from the Missouri Botanical Garden, 93, 1-595.
- Dressler, R. L. & Pupulin, F. (2014). Two new white-flowered species of Sobralia (Orchidaceae) from Costa Rica. Harvard Papers in Botany, 19(1), 117-120.
- Dressler, R. L. & Pupulin, F. (2015). Sobralia lentiginosa (Orchidaceae: Sobralieae). An attractive new species from Costa Rica. Lindleyana in Orchids (Bull. Amer. Orch. Soc.), 84(6), 374-376.
- Dressler, R. L., Acuña, M. & Pupulin, F. (2016). Sobralia turrialbina (Orchidaceae, Sobralieae): long cultivated and now described. Harvard Papes in Botany, 21(2), 251-261.
- Dressler, R. L., Fernández, M. & Pupulin, F. (2014). Sobralia abel-arayae, a new and scarce species from Costa Rica. Orchid Digest, 78(3), 146-148.
- Escobar, R. & Munera, J. M. (1990). Orquídeas nativas de Colombia. Volumen III. Maxillaria-Ponthieva Colina, Medellín, Colombia: Sociedad Colombiana de Orquideología.
- Fernández, M., Bogarín, D., Karremans, A. P. & Jiménez, D. (2014). New species and records of Orchidaceae from Costa Rica. III. Lankesteriana, 13(3), 259-282.
-
Fernández, M., Bogarín, D. & Pupulin, F. (2021). A new Muscarella (Orchidaceae: Pleurothallidinae) from Tapantí National Park, Costa Rica. Webbia, 76(1), 65-70. doi: 10.36253/jopt-10029
» https://doi.org/10.36253/jopt-10029 - Garay, L. A. & Wirth, M. (1959). On the genera Mormolyca Fenzl and Cyrtoglottis Schltr. Canadian Journal of Botany, 37(3), 479-490.
- Gerlach, G. & Pupulin, F. (2020). Stanhopeinae Mesoamericanae VI. On the identity of Polycycnis barbata (Orchidaceae), and other notes on the genus Polycycnis. Phytotaxa, 432(2), 155-170.
-
Hoskovec, L. (2015). Phalaenopsis stuartiana Retrieved from Retrieved from https://botany.cz/cs/phalaenopsis-stuartiana/
Accessed October 2023.
» https://botany.cz/cs/phalaenopsis-stuartiana/ -
Karremans, A. P. (2021). With great biodiversity comes great responsibility: the underestimated diversity of Epidendrum. (Orchidaceae). Harvard Papers in Botany, 26(2), 299-369. https://doi.org/10.3100/hpib.v26iss2.2021.n1
» https://doi.org/https://doi.org/10.3100/hpib.v26iss2.2021.n1 - Karremans, A. P. (2023). Demystifying Orchid Pollination, Stories of Sex, Lies and Obsession Kew: Royal Botanic Gardens. 442 pp.
- Karremans, A. P. & Bogarín, D. (2017). Two novelties in genus Platystele (Orchidaceae: Pleurothallidinae) from Costa Rica. Lankesteriana, 17(2), 215-221.
-
Karremans, A. P. & Díaz-Morales, M. (2017). Novelties in Costa Rican Stelis (Orchidaceae: Pleurothallidinae): two new species and a new record in the “Dracontia group”. Lankesteriana, 17(2), 193-202. https://doi.org/10.15517/lank.v17i2.29928
» https://doi.org/https://doi.org/10.15517/lank.v17i2.29928 - Karremans, A. P. & Jiménez, J. E. (2018). Pleurothallis hawkingii and Pleurothallis vide-vallis (Orchidaceae; Epidendroideae), two new species from Cordillera de Guanacaste in Costa Rica. Phytotaxa, 349(2), 185-191.
- Karremans, A. P. & Jiménez, J. E. (2023). Pleurothallis pugio (Orchidaceae), a new Pleurothallidinae from Costa Rica. Phytotaxa, 599(3), 150-154.
- Karremans, A. P. & Lehmann, C. P. (2018). A highly threatened new species of Vanilla from Costa Rica. Lindleyana in Orchids (Bull. Amer. Orch. Soc.), 304-307.
- Karremans, A. P. & Pupulin, F. (2022). Scaphosepalum Pfitzer. In: F. Pupulin and collaborators (Eds.), Vanishing Beauty, Native Costa Rican Orchids. 1 ed. Vol. 3. Restrepia-Zootrophion and appendices (pp. 1048-1055). Oberreifenberg: Koeltz Botanical Books.
- Karremans, A. P. & Vieira-Uribe, S. (2020). Pleurothallids Neotropical Jewels - Volume I Quito, Ecuador: Imprenta Mariscal. 320 pp.
- Karremans, A. P., Aguilar-Sandí, D., Artavia-Solís, M., Cedeño-Fonseca, M., Chinchilla, I. F., Gil-Amaya, K., Rojas-Alvarado, G., Solano-Guindon, N. & Villegas-Murillo, J. (2019). Nomenclatural notes in the Pleurothallidinae (Orchidaceae): miscellaneous. Phytotaxa, 406(5), 259-270.
- Karremans, A. P., Albetazzi, F. J., Bakker, F. T., Bogarín, D., Eurlings, M. C. M., Pridgeon, A., Pupulin, F. & Gravendeel, B. (2016). Phylogenetic reassessment of Specklinia and its allied genera in the Pleurothallidinae (Orchidaceae). Phytotaxa, 272(1), 1-36.
- Karremans, A. P., Bogarín, D., Fernández, M., Smith, C. & Blanco, M. A. (2012). New species and records of Orchidaceae from Costa Rica. II. Lankesteriana, 12(1), 19-51.
- Karremans, A. P., Bogarín, D., Pupulin, F., Luer, C. A. & Gravendeel, B. (2015). The glandulous Specklinia: morphological convergence versus phylogenetic divergence. Phytotaxa, 218 (2), 101-127.
- Karremans, A. P., Pupulin, F. & Gravendeel, B. (2015). Specklinia dunstervillei, a New Species Long Confused with Specklinia endotrachys (Orchidaceae: Pleurothallidinae). PLoS ONE, 10(7), e0131971.
- Karremans, A. P., Salguero, G., Bogarín, D., Oses, L. & Cedeño-Fonseca, M. (2020). Illustrations and studies in Neotropical Orchidaceae: the Specklinia brighamii group (Pleurothallidinae) in Costa Rica. Phytotaxa, 447 (1), 16-30.
-
Kolanowska, M. (2014). Notes on Costa Rican Pterichis (Orchidaceae) - new taxa and additions to national orchid flora. Lankesteriana, 14(2), 99-108. Doi: 10.15517/lank.v14i2.15586
» https://doi.org/10.15517/lank.v14i2.15586 - Larsen, B. (2014). Three new species of Lepanthes (Orchidaceae). Phytotaxa, 175 (5), 275-280.
- Lindley, J. (1838). Miscellaneous notices. Edwards Botanical Register, 24 (n.s. 11), 1-95.
- Luer, C. A. (1983). New species of Lepanthes (Orchidaceae). Phytologia, 54(4), 325-378.
- Luer, C. A. (1987). A resolution to the problem of Lepanthes turialvae in Costa Rica and Panama. Lindleyana, 2(4), 180-184.
- Luer, C. A. (1993). Icones Pleurothallidinarum X. Systematics of Dracula (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 46, 1-244.
- Luer, C. A. (1996). Icones Pleurothallidinarum XIV. The genus Lepanthes subgenus Lepanthes in Ecuador (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 61(3), 1-255.
- Luer, C. A. (2001). Miscellaneous new species in the Pleurothallidinae (Orchidaceae). Selbyana, 22(2), 103-127.
- Luer, C. A. (2004). Icones Pleurothallidinarum Vol XXVI. Pleurothallis subgenus Acianthera and Three Allied subgenera, A Second Century of new Species of Stelis of Ecuador, Epibator, Ophidion and Zootrophion Monographs in Systematic Botany from the Missouri Botanical Garden, 95, 1- 265.
- Luer, C. A. (2006). Icones Pleurothallidinarum XXVIII. A reconsideration of Masdevallia, and the Systematics of Specklinia and vegetatively similar genera (orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 105, 1-274.
- Luer, C. A. (2007). Icones Pleurothallidinarum XXIX: A Third Century of Stelis of Ecuador, Systematics of Apoda-Prorepentia. Systematics of Miscellaneous Small Genera (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 1-130.
- Luer, C. A. & Toscano de Brito, A. L. V. (2018). Miscellaneous new species in the Pleurothallidinae (Orchidaceae). Harvard Papers in Botany, 23, 47-51.
- Mansfeld, R. (1929). Figuren-Atlas zu den Orchideenfloren der südamerikanischen Kordillerenstaaten von R. Schlechter. Repertorium Specierum Novarum Regni Vegetabilis, Beihefte, 57, t. 1-142.
-
Mondragón-Palomino, M. & Theißen, G. (2009). Why are orchid flowers so diverse? Reduction of evolutionary constraints by paralogues of class B floral homeotic genes, Annals of Botany, 104(3), 583-594. https://doi.org/10.1093/aob/mcn258
» https://doi.org/https://doi.org/10.1093/aob/mcn258 -
Ormerod, P. (2014). Crossoglossa Dressler and Dodson (Orchidaceae: Malaxideae) - An Addendum. Harvard Papers in Botany, 19(1), 97-115. Doi: 10.3100/hpib.v19iss1.2014.n7
» https://doi.org/10.3100/hpib.v19iss1.2014.n7 - Oses, L. (2017). Filogenia molecular de las especies de Masdevallia Ruiz y Pav. (Orchidaceae: pleurothallidinae) de Costa Rica. Tesis (Licenciatura en biología con énfasis en biología molecular y biotecnología). Universidad de Costa Rica, San José, Costa Rica.
- Pupulin, F. (2010). Family #39, Orchidaceae: Tribe Cymbidieae: Subtribe Zygopetalinae. In: W. Burger (ed.), Flora Costaricensis. Fieldiana, Botany n.s. 49.
- Pupulin, F. (2015). × Cochlezella costaricensis, a name for a rare natural hybrid in the Zygopetalinae (Orchidaceae). Harvard Papers in Botany, 20(2), 199-208.
- Pupulin, F. (2019). Zygopetalinae novae et criticae (Orchidaceae). Harvard Papers in Botany, 24(2), 291-339.
- Pupulin, F. (2021). A correction to the Lepanthes guatemalensis group (Orchidaceae: Pleurothallidinae) in Costa Rica, with a new species. Phytotaxa, 480 (1), 69-78.
- Pupulin, F. & collaborators (2022). Vanishing Beauty. Native Costa RicanOrchids. Vol. 3: Restrepia Zootrophion and Appendices Oberreifenberg, Germany: Koeltz Botanical Books.
- Pupulin, F. & Díaz-Morales, M. (2022). Sobralia Ruiz & Pav. In: F. Pupulin and collaborators (Eds.), Vanishing Beauty, Native Costa Rican Orchids 1 ed. Vol. 3. Restrepia-Zootrophion and appendices (pp. 1085-1109). Oberreifenberg: Koeltz Botanical Books .
- Pupulin, F. & Karremans, A. P. (2020). A new and unusual species of Dichaea (Orchidaceae: Zygopetalinae) from Costa Rica. Blumea, 65, 61-64.
- Pupulin, F. & Rojas-Alvarado, G. (2022). Zootrophion Luer. In: F. Pupulin and collaborators (Eds.), Vanishing Beauty, Native Costa Rican Orchids 1 ed. Vol. 3. Restrepia-Zootrophion and appendices (pp. 1375-1381). Oberreifenberg: Koeltz Botanical Books .
- Pupulin, A., Aguilar†, J., Belfort-Oconitrillo, N., Díaz-Morales, M. & Bogarín, D. (2021). Florae Costaricensis subtribui Pleurothallidinis (Orchidaceae) Prodromus II. Systematics of the Pleurothallis cardiothallis and P. phyllocardia groups, and other related groups of Pleurothallis with large vegetative habit. Harvard Papers in Botany, 26(1), 203-295.
- Pupulin, F., Álvarez-Alcázar, L. & Bogarín, D. (2022). A just overtime discovery: another new species of Echinosepala (Orchidaceae) from Costa Rica. Blumea, 67,109-112.
- Pupulin, F., Bogarín, D. & Karremans, A. P. (2020a). Una nueva especie de Cischweinfia (Orchidaceae: Oncidiinae) de Costa Rica, con hábito y flores muy pequeñas. Orquideología, 37(1), 4-16.
-
Pupulin, F., Bogarín, D. & Karremans, A. P. (2023). The Lankester Catalogue of Costa Rican Orchidaceae. Lankesteriana, 23(Supplement), 1-254. http://dx.doi.org/10.15517/lank.v23iSupplement.58145
» https://doi.org/http://dx.doi.org/10.15517/lank.v23iSupplement.58145 - Pupulin, F., Belfort-Oconitrillo, N., Karremans, A. P. & Bogarín, D. (2020b). Florae Costaricensis Subtribui Pleurothallidinis Prodromus-Systematics of Echinosepala (Orchidaceae). Harvard Papers in Botany, 25(2), 155-190.
- Pupulin, F., Chinchilla, I. F., Rojas-Alvarado, G., Fernández, M., Ossenbach, C. & Bogarín, D. (2022). Typification of Costa Rican Orchidaceae described by Rudolf Schlechter. Species variorum collectorum. Webbia, 77(1), 35-126.
- Pupulin, F., Díaz-Morales, M., Aguilar, J. & Fernández, M. (2017a). Two new species of Pleurothallis (Orchidaceae: Pleurothallidinae) allied to P. cardiothallis, with a note on flower activity. Lankesteriana, 17(2), 329-356.
- Pupulin, F., Díaz-Morales, M., Fernández, M. & Aguilar, J. (2017b). Two new species of Pleurothallis (Orchidaceae: Pleurothallidinae) from Costa Rica in the P. phyllocardia group. Lankesteriana, 17(2), 153-164.
- Pupulin, F., Karremans, A. P. & Belfort-Oconitrillo, N. (2017c). Two new species of Echinosepala (Orchidaceae: Pleurothallidinae). Lankesteriana, 17(2), 285-304.
- Pupulin, F., Merino, G. & Medina, H. (2009). Draculas del Ecuador. Quito. Ecuador: Editorial Universidad Alfredo Pérez Guerrero.
- Rojas-Alvarado, G. A. (2023). A new species of Stelis subgen. Unciferia (Orchidaceae: Pleurothallidinae) from Costa Rica. Phytotaxa, 607(3), 205-212.
- Rojas-Alvarado, G. A. & Karremans, A. P. (2017). Additions to the costa rican Myoxanthus (Orchidaceae: Pleurothallidinae). Lankesteriana, 17(2), 203-214.
- Rojas-Alvarado, G. A. & Karremans, A. P. (2020). Revision of the Costa Rican species of Myoxanthus (Pleurothallidinae: Orchidaceae). Phytotaxa, 448(1), 1-70.
- Rojas-Alvarado, G. A., Karremans, A. P. & Blanco, M. A. (2021). A taxonomic synopsis and morphological characterization of Myoxanthus (Orchidaceae: Pleurothallidinae). Phytotaxa 507(3), 211-258.
- Schlechter, R. (1917). Neue und seltene Garten-Orchideen. Orchis. Monatsschrift der Deutschen Gesellschaft für Orchideenkunde, 6(7), 112-119.
- Smith, C. M., Bogarín, D. & Pupulin, F. (2015). A new Reichantha (Orchidaceae: Pleurothallidinae) from Nicaragua and Costa Rica. Systematic Botany, 40(1), 83-87.
- Soto Arenas, M., Salazar, G. A. & Solano, R. (2002). Dryadella greenwoodiana In: E. Hágsater & M. Soto (Ed.), Icones Orchidacearum 5-6 (t. 550). México D.F.: Instituto Chinoin, A. C.
- Soto Arenas, M. A. & Solano, R. (2003). Specklinia echinata In: E. Hágsater & M. A. Soto Arenas (eds.), Icones Orchidacearum (Mexico), Fascicles 5 and 6. Orchids of Mexico Parts 2 and 3. Herbario AMO. México D.F.: Instituto Chinoin, A.C. pl. 670.
- Schweinfurth, C. (1944). Mormolyca peruviana American Orchid Society Bulletin, 13, 196.
- Schweinfurth, C. (1960). Orchidaceae, Orchids of Peru. Fieldiana, Botany, 30(3), 533-786.
- Szlachetko, D. L., Sitko, M., Tukallo, P. & Mytnik-Ejsmont, J. (2012). Taxonomy of the subtribe Maxillariinae (Orchidaceae, Vandoideae) revised. Biodiversity Research and Conservation, 25, 13-38.
-
Thiers, B. (2023). Index herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. Retrieved from Retrieved from http://sweetgum.nybg.org/science/ih/
[accessed 1 Jan 2023]
» http://sweetgum.nybg.org/science/ih/ - Vásquez C., R. & Dodson, C. H. (1982). Orchids of Bolivia. Icones Plantarum Tropicarum, 6, 501-600.
- Vieira-Uribe, S. & Bogarín, D. (2016). Una hermosa nueva especie de Masdevallia (Pleurothallidinae: Orchidaceae) de los Andes centrales de Colombia. Orquideología, 33, 14-19.
Publication Dates
-
Date of issue
May-Aug 2024
History
-
Received
20 Nov 2023 -
Accepted
18 June 2024

 LCDP prepared by G. Rojas-Alvarado based on D.Bogarín 10298. A. Habit. B. Flower, – view. C. Perianth dissected. D. Column with lip, lateral view. E. Lip, – view. F. Column, – and ventral views. G. Anther cap and pollinarium.
LCDP prepared by G. Rojas-Alvarado based on D.Bogarín 10298. A. Habit. B. Flower, – view. C. Perianth dissected. D. Column with lip, lateral view. E. Lip, – view. F. Column, – and ventral views. G. Anther cap and pollinarium.
 Photos by K. Gil-Amaya based on N.Belfort-Oconitrillo 380. LCDP prepared by K. Gil-Amaya and N. Belfort-Oconitrillo. A. Habit. B. Flower. C. Dissected perianth. D. Column and lip in natural position, lateral view. E. Lip, ventral and dorsal views. F. Petal, ventral, dorsal and lateral views. G. Column, ventral and – views. H. Anther cap and pollinarium.
Photos by K. Gil-Amaya based on N.Belfort-Oconitrillo 380. LCDP prepared by K. Gil-Amaya and N. Belfort-Oconitrillo. A. Habit. B. Flower. C. Dissected perianth. D. Column and lip in natural position, lateral view. E. Lip, ventral and dorsal views. F. Petal, ventral, dorsal and lateral views. G. Column, ventral and – views. H. Anther cap and pollinarium.
 Drawn by D. Bogarín & L. Oses based on D.Bogarín 11108 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Ovary, column and lip, lateral view. E. Lip, flattened. F. Column, ventral view. G. Anther cap and pollinarium.
Drawn by D. Bogarín & L. Oses based on D.Bogarín 11108 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Ovary, column and lip, lateral view. E. Lip, flattened. F. Column, ventral view. G. Anther cap and pollinarium.
 Photos by D. Bogarín (A) and G. Salguero (B).
Photos by D. Bogarín (A) and G. Salguero (B).
 LCDP prepared by G. Salguero based on A.P.Karremans 6720. A. Habit. B. Inflorescence. C. Flower. D. Dissected perianth. E. Lip and column in natural position, lateral view. F. Column in lateral, – and ventral views. G. Anther cap and pollinarium.
LCDP prepared by G. Salguero based on A.P.Karremans 6720. A. Habit. B. Inflorescence. C. Flower. D. Dissected perianth. E. Lip and column in natural position, lateral view. F. Column in lateral, – and ventral views. G. Anther cap and pollinarium.
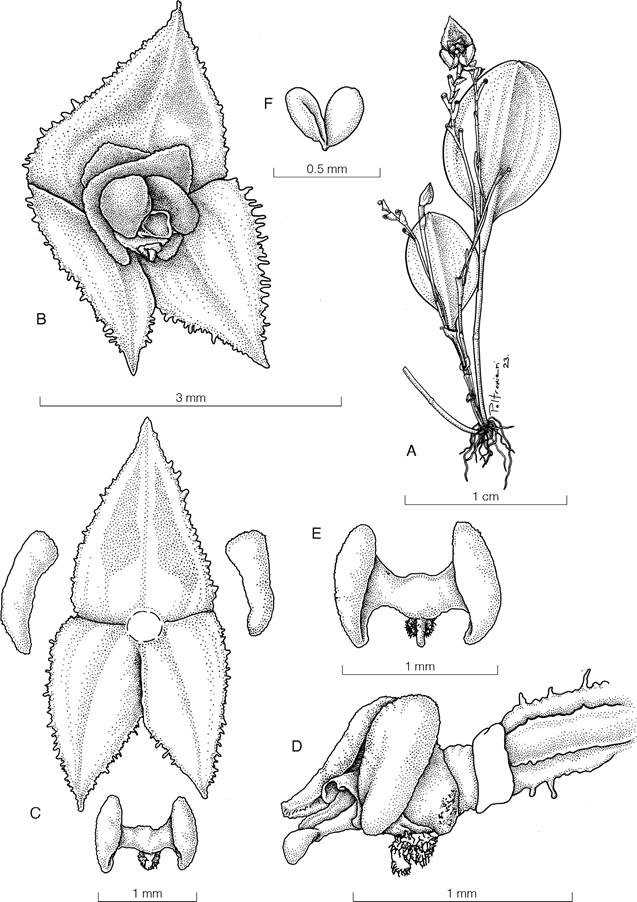 Drawn by D. Bogarín, K. Gil-Amaya and S. Poltronieri based on D.Bogarín 12501 (JBL-spirit). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, lateral view. E. Lip, spread. F. Pollinia.
Drawn by D. Bogarín, K. Gil-Amaya and S. Poltronieri based on D.Bogarín 12501 (JBL-spirit). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, lateral view. E. Lip, spread. F. Pollinia.
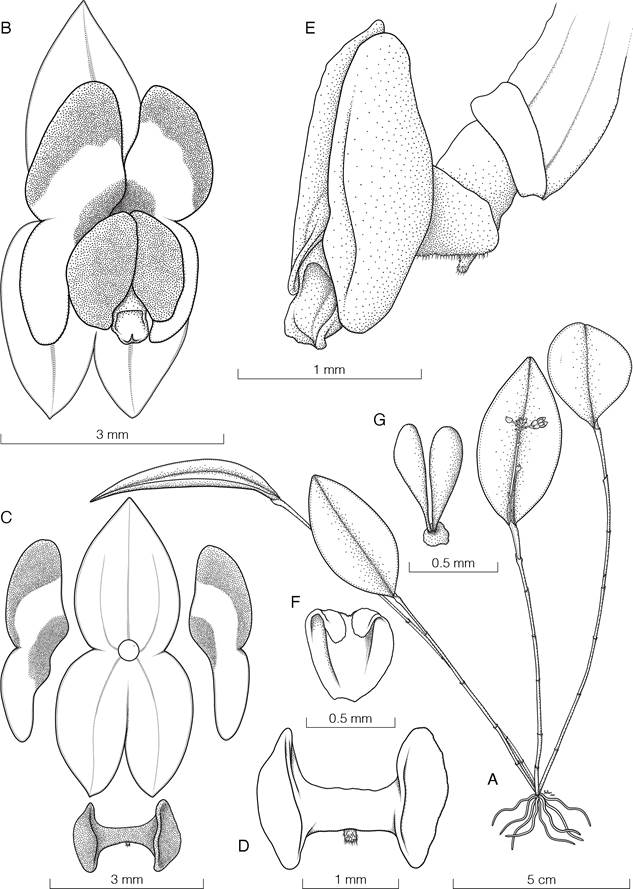 Drawn by I. F. Chinchilla and D. Bogarín from D.Bogarín 8243 (JBL spirit). A. Habit. B. Flower, in natural position. C. Dissected perianth, flattened. D. Close up of lip spread, adaxial view. E. Ovary, column and lip, oblique view. F. Anther cap, abaxial view. G. Pollinarium.
Drawn by I. F. Chinchilla and D. Bogarín from D.Bogarín 8243 (JBL spirit). A. Habit. B. Flower, in natural position. C. Dissected perianth, flattened. D. Close up of lip spread, adaxial view. E. Ovary, column and lip, oblique view. F. Anther cap, abaxial view. G. Pollinarium.
 Photographs by D. Bogarín.
Photographs by D. Bogarín.
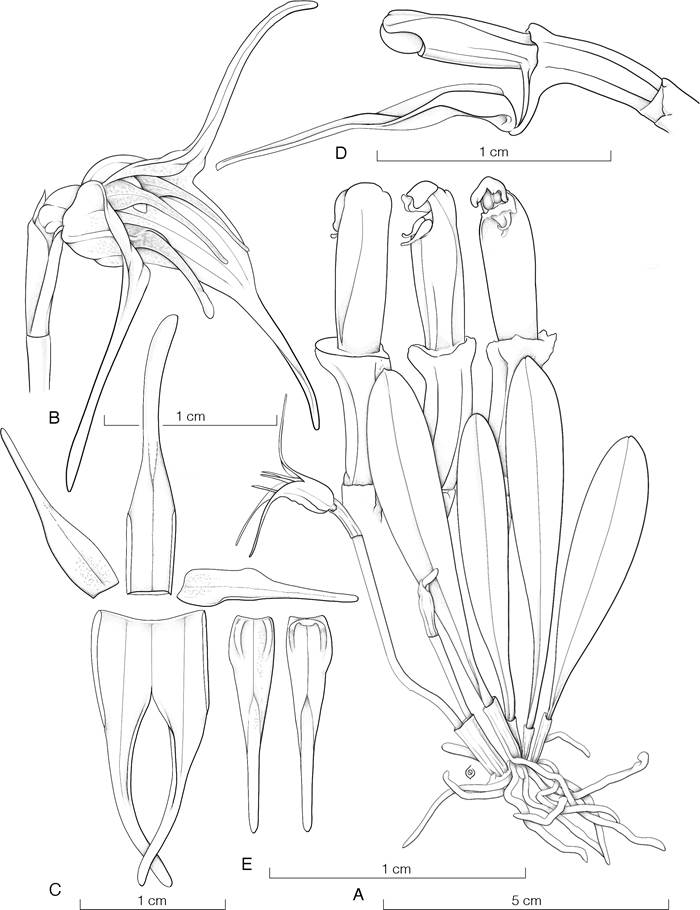 Drawn by L.Oses based on D.Bogarín 10767 (JBL-spirit). A. Habit. B. Flower. C. Perianth with ventral and dorsal view of the lip. D. Column, lip, and ovary lateral view. E. Column, ventral view.
Drawn by L.Oses based on D.Bogarín 10767 (JBL-spirit). A. Habit. B. Flower. C. Perianth with ventral and dorsal view of the lip. D. Column, lip, and ovary lateral view. E. Column, ventral view.
 LCDP prepared by N. Belfort-Oconitrillo based on N.Belfort-Oconitrillo 938 (JBL-spirit). A. Habit. B. Flower. C. Dissected perianth. D. Ovary, column and lip, lateral view. E. Lip, adaxial and abaxial views. F. Column, – view. G. Anther cap and pollinarium.
LCDP prepared by N. Belfort-Oconitrillo based on N.Belfort-Oconitrillo 938 (JBL-spirit). A. Habit. B. Flower. C. Dissected perianth. D. Ovary, column and lip, lateral view. E. Lip, adaxial and abaxial views. F. Column, – view. G. Anther cap and pollinarium.
 Plate prepared by N. Belfort-Oconitrillo based N.Belfort-Oconitrillo 676 (JBL-spirit). A. Flower. B. Dissected perianth. C. Ovary, column and lip, lateral view. D. Lip in adaxial, –, lateral, and abaxial views. E. Column. F. Anther cap. G. Pollinarium.
Plate prepared by N. Belfort-Oconitrillo based N.Belfort-Oconitrillo 676 (JBL-spirit). A. Flower. B. Dissected perianth. C. Ovary, column and lip, lateral view. D. Lip in adaxial, –, lateral, and abaxial views. E. Column. F. Anther cap. G. Pollinarium.

 LCDP prepared by F. Pupulin based on L.Álvarez 499 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, lateral view. E. Lip in longitudinal section. F. Column in several views. G. Anther cap. H. pollinarium in dorsal, ventral, and lateral views.
LCDP prepared by F. Pupulin based on L.Álvarez 499 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, lateral view. E. Lip in longitudinal section. F. Column in several views. G. Anther cap. H. pollinarium in dorsal, ventral, and lateral views.
 Vouchers at JBL. Photographs by F. Pupulin. A. Álvarez 335.
B. Álvarez 1229.
Vouchers at JBL. Photographs by F. Pupulin. A. Álvarez 335.
B. Álvarez 1229.
 All vouchers at JBL. Digital composite plate prepared by F. Pupulin. A. M. cryptobulbon (A.P.Karremans 6622). B. M. ringens (D.Bogarín 11479). C. M. angustisegmenta (F.Pupulin 461). D. M. brunnea (D.Bogarín 1575). E. M. porrecta (A.P.Karremans 6727). F. M. hennisiana (L.Álvarez 355). Scale bar = 3 cm.
All vouchers at JBL. Digital composite plate prepared by F. Pupulin. A. M. cryptobulbon (A.P.Karremans 6622). B. M. ringens (D.Bogarín 11479). C. M. angustisegmenta (F.Pupulin 461). D. M. brunnea (D.Bogarín 1575). E. M. porrecta (A.P.Karremans 6727). F. M. hennisiana (L.Álvarez 355). Scale bar = 3 cm.
 LCDP prepared by F. Pupulin based on A.P.Karremans 6671 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Lip and column in natural position, lateral view. E. Lip, longitudinal section. F. Column, – and ventral views (emasculate on the left). G. Pollinarium in abaxial, adaxial and lateral views, and anther cap.
LCDP prepared by F. Pupulin based on A.P.Karremans 6671 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Lip and column in natural position, lateral view. E. Lip, longitudinal section. F. Column, – and ventral views (emasculate on the left). G. Pollinarium in abaxial, adaxial and lateral views, and anther cap.
 LCDP prepared by D. Bogarín and F. Pupulin based on D.Bogarín 13092 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Lip in lateral, abaxial, and adaxial views. E. Lip and column in natural position, lateral view. F. Column in lateral and ventral views. G. Anther cap. H. Pollinarium in abaxial, adaxial and lateral views.
LCDP prepared by D. Bogarín and F. Pupulin based on D.Bogarín 13092 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Lip in lateral, abaxial, and adaxial views. E. Lip and column in natural position, lateral view. F. Column in lateral and ventral views. G. Anther cap. H. Pollinarium in abaxial, adaxial and lateral views.
 LCDP prepared by F. Pupulin based on F.Pupulin 9082 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, – view. E. Column, ventral view (emasculate on the right). F. Anther cap. G. pollinarium, dorsal view. H. Pollinarium, – and ventral views.
LCDP prepared by F. Pupulin based on F.Pupulin 9082 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, – view. E. Column, ventral view (emasculate on the right). F. Anther cap. G. pollinarium, dorsal view. H. Pollinarium, – and ventral views.
 Photograph by Jindřiška Vančurová.
Photograph by Jindřiška Vančurová.
 LCDP prepared by F. Pupulin based on D.Bogarín 8427 (JBL). A. Habit. B. Flower. C. Dissected perianth (the lip in ventral, dorsal, and lateral views). D. Column and lip, lateral view. E. Column in lateral, dorsal, and ventral views. F. Anther cap and pollinarium in dorsal and ventral views. G. Pollinarium in three lateral views.
LCDP prepared by F. Pupulin based on D.Bogarín 8427 (JBL). A. Habit. B. Flower. C. Dissected perianth (the lip in ventral, dorsal, and lateral views). D. Column and lip, lateral view. E. Column in lateral, dorsal, and ventral views. F. Anther cap and pollinarium in dorsal and ventral views. G. Pollinarium in three lateral views.
 All the vouchers at JBL. Photographs by F. Pupulin. A. A.P.Karremans 6077. B. Díaz 237. C. P. trigyna (D.Bogarín 8427). D. D.Bogarín 9091. E. D.Bogarín 8739. Scale bar = 5 mm.
All the vouchers at JBL. Photographs by F. Pupulin. A. A.P.Karremans 6077. B. Díaz 237. C. P. trigyna (D.Bogarín 8427). D. D.Bogarín 9091. E. D.Bogarín 8739. Scale bar = 5 mm.
 Photomacrography by F. Pupulin based on D.Bogarín 8427.
Photomacrography by F. Pupulin based on D.Bogarín 8427.
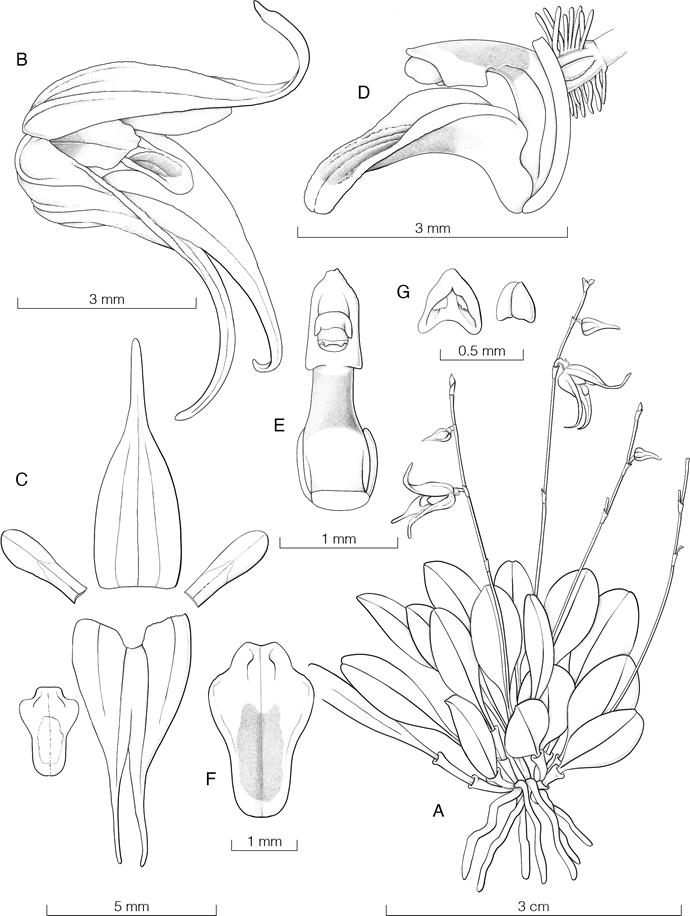 Drawn by D. Bogarín and L.Oses based on A.P.Karremans 5600. A. Habit. B. Flower. C. Dissected perianth. D. Ovary, column and lip, lateral view. E. Column, ventral view. F. Lip. G. Anther cap and pollinia.
Drawn by D. Bogarín and L.Oses based on A.P.Karremans 5600. A. Habit. B. Flower. C. Dissected perianth. D. Ovary, column and lip, lateral view. E. Column, ventral view. F. Lip. G. Anther cap and pollinia.
 LCDP prepared by A.P. Karremans based on A.P.Karremans 8823 (A) and A.P.Karremans 8822 (B-E). A. Habit with three inflorescences. B. Habit with an open flower. C. Dissected perianth. D. Column and ovary in ventral and lateral views. E. Anther cap and pollinia.
LCDP prepared by A.P. Karremans based on A.P.Karremans 8823 (A) and A.P.Karremans 8822 (B-E). A. Habit with three inflorescences. B. Habit with an open flower. C. Dissected perianth. D. Column and ovary in ventral and lateral views. E. Anther cap and pollinia.
 Drawn by I. F. Chinchilla from D.Bogarín 10490 (JBL-spirit). A. Habit. B. Portion of inflorescences showing three flowers. C. Flower, in natural position. D. Sepals, petals, lip and column, frontal view.
Drawn by I. F. Chinchilla from D.Bogarín 10490 (JBL-spirit). A. Habit. B. Portion of inflorescences showing three flowers. C. Flower, in natural position. D. Sepals, petals, lip and column, frontal view.
 LCDP prepared by I. F. Chinchilla from D.Bogarín 10490 (JBL-spirit). A. Habit. B. Portion of inflorescences showing three flowers. C. Flower, in natural position. D. Sepals, petals, lip and column, frontal view.
LCDP prepared by I. F. Chinchilla from D.Bogarín 10490 (JBL-spirit). A. Habit. B. Portion of inflorescences showing three flowers. C. Flower, in natural position. D. Sepals, petals, lip and column, frontal view.
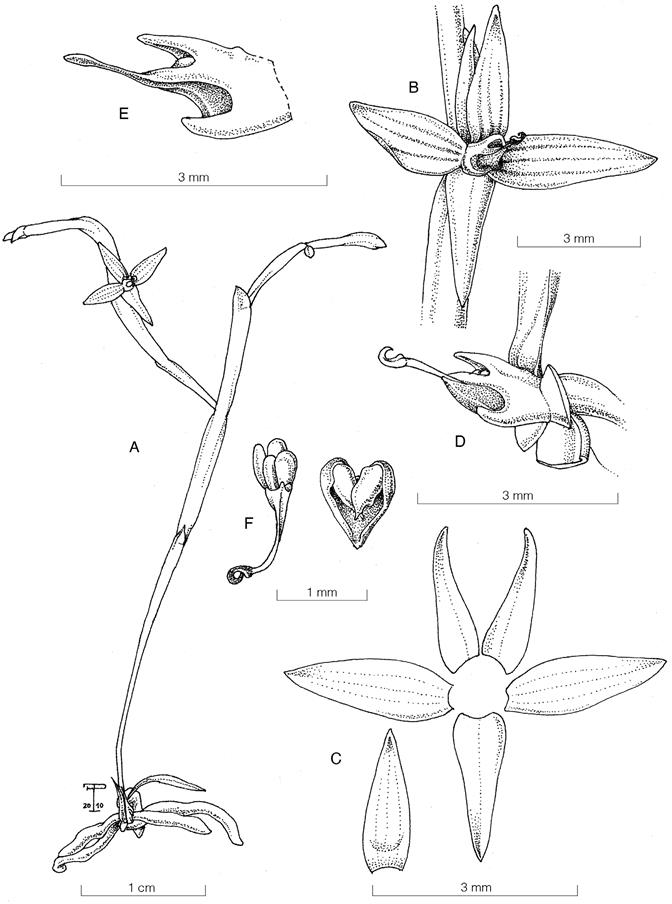 Drawn by F. Pupulin based on F.Pupulin 3557 (USJ). A. Habit. B. Flower. C. Dissected perianth. D. Column, lateral view, showing the bases of sepals, petals, and lip. E. Column in lateral view, emasculate. F. Anther cap and pollinarium.
Drawn by F. Pupulin based on F.Pupulin 3557 (USJ). A. Habit. B. Flower. C. Dissected perianth. D. Column, lateral view, showing the bases of sepals, petals, and lip. E. Column in lateral view, emasculate. F. Anther cap and pollinarium.
 Vouchers: F.Pupulin et al. 3557 (A), D.Bogarín et al. 6582 (B), and D.Bogarín 13425 & Villalobos (C).
Vouchers: F.Pupulin et al. 3557 (A), D.Bogarín et al. 6582 (B), and D.Bogarín 13425 & Villalobos (C).
 LCDP prepared by F. Pupulin based on F.Pupulin 9118 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, lateral view (the lip in longitudinal section). E. Lip section. F. Column in several views. G. Anther cap. H. Pollinarium in dorsal, ventral, and lateral views.
LCDP prepared by F. Pupulin based on F.Pupulin 9118 (JBL). A. Habit. B. Flower. C. Dissected perianth. D. Column and lip, lateral view (the lip in longitudinal section). E. Lip section. F. Column in several views. G. Anther cap. H. Pollinarium in dorsal, ventral, and lateral views.